
Разнообразие смерти: гастрономические предпочтения змей и эволюция их яда

В мире людей существует немало страхов, некоторые их которых перерастают в самые настоящие фобии: арахнофобия (пауки), акрофобия (высота), аквафобия (вода), коулрофобия (клоуны), офидиофобия (змеи) и т.д. Многие люди, даже не имея офидиофобии, относятся к змеям с призрением и недоверием. Змей часто отожествляют с хитростью, подлостью, коварством и прочими малоприятными качествами, которые по иронии им совершенно неприсущи, в отличие от самих людей. С другой стороны змеи часто символизируют мудрость, бессмертие, знания. Одними из самых узнаваемых символов, в которых присутствует змея, являются сосуд Гигиеи (символ фармации) и посох Асклепия (символ медицины). Для многих самой примечательной особенностью некоторых змей является их яд, структура которого может быть невероятна. Ученые из Клемсонского университета (США) выяснили, что структура яда некоторых змей зависит от того, насколько эволюционно далека друг от друга добыча. Как гастрономические предпочтения змей влияют на их яд, чей яд сильнее, и как полученные знания можно применить на практике? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Многие виды на нашей планете связаны друг с другом. Это вполне логично, ибо есть хищники, поедающие добычу, есть паразиты, не способные жить без носителя, есть симбиотические существа, получающие выгоду от совместного проживания с другими видами. Связь проявляется по-разному, но она одинаково играет важную роль в развитии каждого из участников этой связи. Об этом говорил еще Дарвин, уделяющий внимание в своих трудах корреляции между эволюцией признаков и взаимодействиями видов.
Однако связь не формируется сразу, на это требуется уйма времени, в течение которого происходит процесс естественного отбора. Ученые приводят в пример связь носителя и паразитов. Носитель в ходе эволюции может развить уникальную для каждого вида паразита защиту или же универсальную — сразу от нескольких видов паразитов. В таком случае необходимо понять, важно ли количество или все же характеристики взаимодействующих видов.
Сложные черты того или иного вида определяются множеством компонентов, вносящих вклад в окончательный функциональный фенотип, который формируется в зависимости от количества информации в геноме, определенной данными компонентами. Молекулярные признаки, участвующие в антагонистических взаимодействиях, проявляются как модели для связывания сложности признака с разнообразием видовых сообществ, поскольку их сложность может быть точно определена количественно числом и разнообразием уникальных компонентов. Это, по словам ученых, позволяет использовать меры разнообразия, такие как H-индекс Шеннона*, для суммирования сложности той или иной черты вида.
Индекс (или энтропия) Шеннона* был предложен Клодом Шенноном в 1948 году для количественной оценки энтропии в строках текста. Суть модели состоит в том, что чем больше разных букв и чем больше их пропорциональное количество в интересующей строке, тем труднее правильно предсказать, какая буква будет следующей в строке. Энтропия Шеннона количественно определяет неопределенность (энтропию), связанную с этим предсказанием.
Одним из ярчайших примеров взаимодействия видов является использование одним их низ яда для защиты/нападения. Яд нарушает гомеостатические физиологические процессы, быстро делая добычу неспособной сопротивляться либо нападающего неспособным продолжить преследование. Это взаимодействие позволяет понять силы, опосредующие сложность признаков во взаимодействиях хищник-жертва.
Что касается белковых смесей, сложность фенотипа яда может быть определена количественно с помощью хроматографии*.
Хроматография* — метод разделения и анализа смесей веществ, а также изучения их физико-химических свойств. Суть метода заключается в распределении вещества между двумя фазами: неподвижная (твердая или жидкая) и подвижная (газовая или жидкая).
Кроме того, транскриптом ядовитой железы напрямую связывает протеом (совокупность белков) яда с генотипом вида. То есть транскриптом является вторым независимым методом определения сложности яда за счет оценки сложности геномной последовательности.
Функциональные исследования змеиного яда выявили несколько примеров разнообразия и специфичности добычи, вызванных эволюционными процессами. К примеру, из-за того, что морские змеи вида Aipysurus eydouxii начали питаться практически исключительно икрой рыб, их яд стал намного слабее, а также были выявлены нулевые и негативные мутации в генах яда.
Любопытно и то, что паралоги* высокоэкспрессированных генов яда одной и той же змеи могут обладать таксономической специфичностью.
Паралоги* — последовательности, которые присутствуют в одном и том же геноме, а возникают в эволюции путем дупликации первичной последовательности.
К примеру, яд змей Spilotes sulphureus одинаково смертелен для млекопитающих или ящериц, а яд Bothrops neuwiedi по-разному выполняет прокоагулянтную функцию (свертывание крови) в крови любого из представителей млекопитающих.
В данном труде особое внимание было уделено гадюковым змеям, так как их гастрономические предпочтения были хорошо изучены, что позволяет детально оценить различия в рационе питания и влияние этого фактора на структуру яда.
Большинство гадюк питаются в основном мелкими позвоночными, при этом диета их предков, вероятно, состояла из млекопитающих, ящериц и лягушек. Гремучие змеи (Crotalus и Sistrurus), медноголовые и мокассиновые (Agkistrodon) составляют крупнейшую кладу переднезубых ядовитых змей в Северной Америке, насчитывающую от 45 до 64 описанных видов. Питание этих видов также обладает широким разнообразием. К примеру, Crotalus horridus питается исключительно млекопитающими, а вот Agkistrodon piscivorus conanti не брезгует рыбой, лягушками, млекопитающими, ящерицами, птицами, черепахами и даже змеями.
Яды этих родов змей состоят из 10–70 белков из 15–25 различных семейств генов. Из этого и происходит разнообразие действия яда: нейротоксическое (нарушение периферической нервной системы), коагулопатическое (нарушение свертывания крови), геморрагическое (образование кровяных сгустков) или миотоксическое (предотвращение релаксации мышц после сокращения) и т.д.
Дабы проверить, какие теории о связи между питанием змей и структурой их яда все же верны, а какие нет, ученые проанализировали рацион змей и измерили протеомную сложность яда и транскриптомную сложность ядовитых желез. Образцы яда и желез были получены от змей Agkistrodon, Crotalus и Sistrurus. В результате была сформирована самая крупная база данных протеомов и транскриптомов ядовитых желез для этой группы на сегодняшний день (68 линий). Была также создана филогения* из 125 неядерных генетических локусов* из транскриптомов, после чего был проведен сопоставительный анализ с рационом змей.
Филогения* — результат филогенетического анализа, т.е. процесса выявления и объяснения эволюционных взаимоотношений между видами.
Локус* — местоположение определенного гена на генетической или цитогенетической карте хромосомы.
Результаты исследования
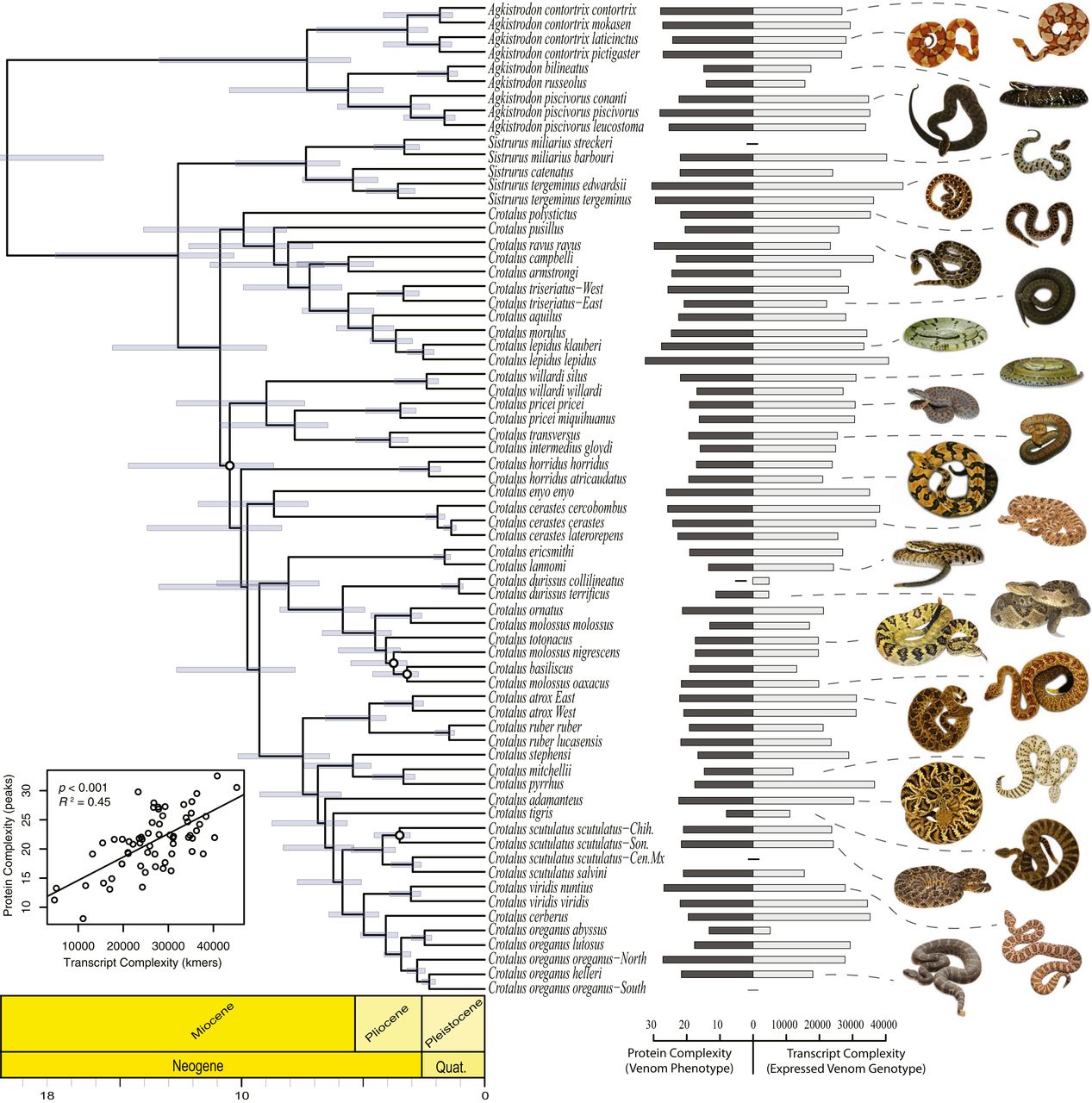
Изображение №1
В ходе исследования была получена филогения посредством методов видового древа (схема выше) и конкатенации* с использованием последовательностей от 169 особей, представляющих 46 видов Agkistrodon, Crotalus и Sistrurus.
Конкатенация* — объединение объектов линейной структуры в одну (например, слова «микро» и «мир» дают «микромир»). В генетике подход, основанный на конкатенации, представляет собой подход «полного доказательства», который объединяет все сопоставления генов в суперматрицу.
Чтобы получить максимальное разнообразие в пределах исследуемой основной клады, выборка была расширена. В нее дополнительно были включены филогенетически отдельные подвиды и линии с описанным филогенетическим разнообразием, что привело к окончательному набору данных о 9 линиях Agkistrodon, 5 линиях Sistrurus и 54 линиях Crotalus.
Полученные деревья были использованы для филогенетического сравнительного анализа взаимосвязи между разнообразием рациона и сложностью яда.
Первый анализ, во время которого оценивалась связь между транскриптомной и протеомной сложностью яда, показал, что между выраженным генотипом и сложностью фенотипа в ядах змей присутствует сильная связь (диаграмма на изображении №1).
Ученые отмечают, что в ходе высокоэффективной жидкостной хроматографии (ВЭЖХ) похожие белки могут элюироваться (извлекаться) вместе в виде одного пика на хроматограммах, что потенциально недооценивает сложность фенотипа на уровне отдельных компонентов. Однако применение меры транскриптомной сложности позволяет количественно определять различия между белками, в то время как равный вес всех уникальных последовательностей независимо от длины транскрипта обеспечивает дополнительный акцент на общем разнообразии последовательностей, а не на организации в транскрипты.
Транскриптомная сложность, лежащая в основе ядов, варьировалась в 10 раз между линиями. При этом самый сложный транскриптом был у змей вида Sistrurus tergeminus edwardsii — 45191 k-мер (последовательностей из k нуклеотидов); самый же простой транскриптом принадлежал виду Crotalus durissus terrificus — 4723 k-мер.
Протеомная сложность яда также значительно различалась: наиболее сложная у Crotalus lepidus lepidus — примерно 32 пика белка на хроматограммах; самая простая у Crotalus tigris — в среднем 8 пиков.
Реконструкции состояния предков указывают на промежуточный белок (22.3 пика) и транскриптомную сложность (28751 k-мер) у общего предка агкистродонов и гремучих змей. Из этого можно сделать вывод, что с течением времени произошла эволюция как в сторону более простых, так и более сложных ядов.
Разнообразие у змей проявляется не только в аспекте видов, но и в аспекте потребляемой пищи (схема ниже), т.е. видового разнообразия добычи, которое варьировалось от 2.9 до 15 (среднее 6.7) видов и от 118 до 731 (среднее значение = 365) MPD (от million years of divergence, т.е. миллионов лет расхождения*).
Дивергенция (расхождение)* — расхождение признаков и свойств у первоначально близких групп организмов в ходе эволюции.
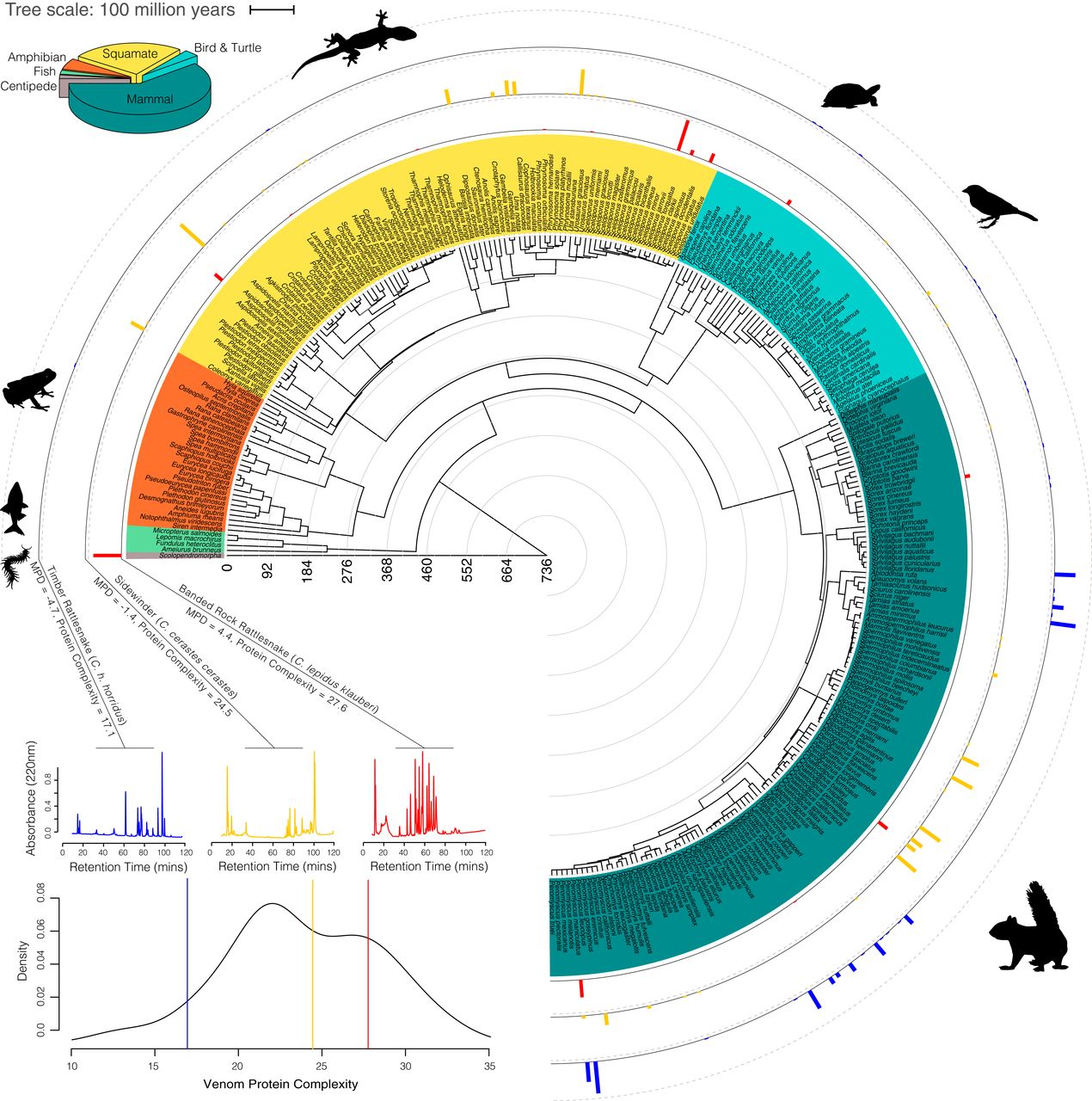
Изображение №2
Наблюдаемые вариации протеомной и транскриптомной сложности яда лучше всего моделировались с помощью MPD добычи по всем альтернативным филогенетическим деревьям, используемым для представления этой взаимосвязи. Кроме того, модели MPD значительно лучше соответствовали данным по сравнению с нулевыми филогенетическими обобщенными моделями наименьших квадратов. Другими словами, используемая модель по MPD показала, что эволюция сама по себе не может объяснить вариативность ядов у змей.
Наряду с положительной корреляцией между сложностью яда и MPD (график ниже), предложенная модель подкрепляет теорию филогенетического разнообразия для эволюции более сложных признаков яда у змей.
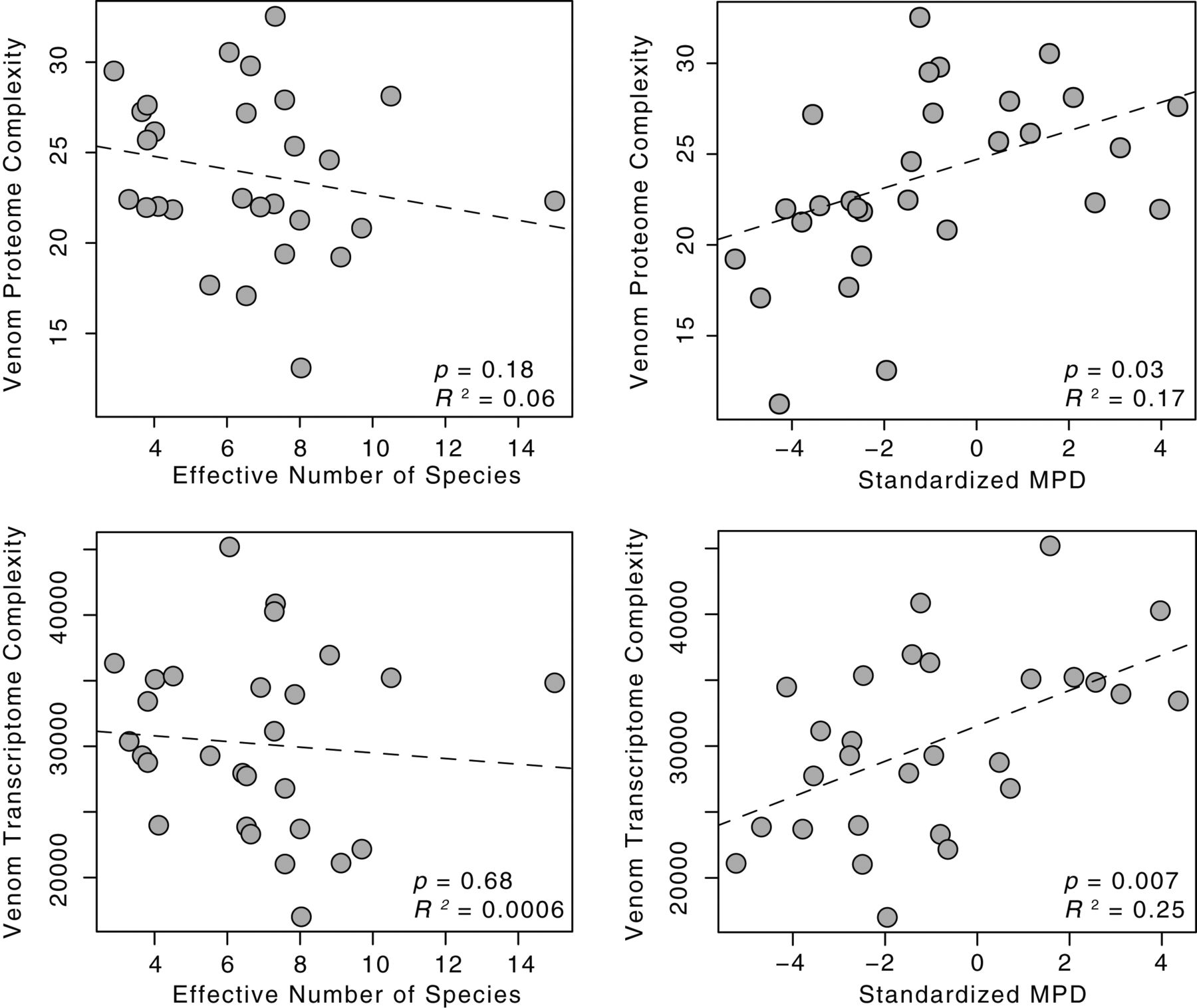
Изображение №3
Более детальный анализ показал положительную корреляцию между сложностью яда и MPD добычи, однако не было выявлено связи между видовым разнообразием и сложностью белков яда. Подобная ситуация наблюдалась и при анализе MPD и транскриптомной сложности: корреляция между ними была, но значимой связи между видовым разнообразием и транскриптомной сложностью яда не было.
Далее была выполнена оценка взаимосвязи между транскриптомной сложностью яда и филогенетическим разнообразием добычи на уровне семейства генов для четырех крупнейших семейств генов среди гадюк (SVMP, SVSP, PLA2 и CTL). Значимые положительные корреляции между сложностью транскриптомов и MPD добычи были обнаружены для семейства генов SVMP, SVSP и PLA2 (график ниже). Семейство генов CTL, однако, не показало значимой связи между MPD и сложностью транскриптомики.

Изображение №4
Данный анализ показывает эволюционный ответ на отбор из более филогенетически разнообразной добычи, происходящий через несколько семейств генов.
В заключение была выполнена проверка того, отражает ли филогенетическое разнообразие добычи функциональное разнообразие яда лучше, чем обычное видовое разнообразие.
Анализ аминокислотных последовательностей яда разных змей показал, что MPD добычи в большей степени отражает разнообразие белковых последовательностей в функционально важных элементах яда, чем простое разнообразие видов добычи. Кроме того, разнообразие аминокислотных последовательностей в конкретных аспектах яда может успешно предсказать его сложность.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Эволюцию сложно назвать быстрым и легким процессом, о каком бы виде не шла речь. Каждый живой организм адаптируется к ряду факторов, в числе которых присутствует и окружающая среда, и ее обитатели. Подавляющее большинство видов так или иначе связано друг с другом. Одни являются хищниками, другие — добычей, одни — паразитами, другие — носителями и т.д. Даже обычное проживание на одной и той же территории видов, которые не контактируют напрямую, вносит свой вклад в их эволюционное развитие.
В данном труде ученые установили, что разнообразие рациона змей коррелирует со сложностью их яда. Другими словами, чем разнообразнее гастрономические аппетиты змеи, тем сложнее будет ее яд. На первый взгляд это кажется весьма очевидным выводом. Далеко не все яды одинаково действуют на разных существ. Следовательно, если есть желание лопать всех подряд, яд должен быть сложным и, следовательно, универсально смертоносным.
Несмотря на простоту вопроса, ответ оказался гораздо сложнее и затронул миллионы лет эволюции. При этом было установлено, что среди четырех семейств генов лишь троим можно приписывать связь видового разнообразия добычи и сложности яда. Четвертое семейство (CTL, т.е. лектины типа С) пошли по другому пути. В дальнейшем ученые намерены выяснить, почему имело место именно такое генное распределение обязанностей.
Важность этого исследования заключается в том, что змеиный яд крайне важный элемент многих препаратов, нацеленных на лечение заболеваний сердца, высокого давления, тромбов и т.д. Змей очень много (порядка 3631 видов по данным 2017 года), среди которых разнообразие ядов еще больше. Чем лучше мы понимаем структуру змеиного яд, тем лучше мы сможем его использовать в фармации.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Похожие статьи
 Как образуются нейтронные звёзды?
Как образуются нейтронные звёзды? Эволюция Microsoft Word: от истоков до современных технологий
Эволюция Microsoft Word: от истоков до современных технологий Тысяча и одна задача для устного счета
Тысяча и одна задача для устного счета Какова мощь электромагнитных сил?
Какова мощь электромагнитных сил? Казань в движении: от телеги водовоза до робота-доставщика
Казань в движении: от телеги водовоза до робота-доставщика Horsey Horseless: почему самый странный автомобиль в истории важен для UI-дизайна
Horsey Horseless: почему самый странный автомобиль в истории важен для UI-дизайна Резервуарные вычисления: как работает нейросеть, которую почти не учат
Резервуарные вычисления: как работает нейросеть, которую почти не учат Подземные города: от древности до недалекого будущего
Подземные города: от древности до недалекого будущего