
В мире дикой природы роли распределены очень просто, либо ты — хищник, либо добыча. За долгие годы эволюции роли эти никуда не делись, но вот формат их реализации претерпел немало изменений в каждом отдельном виде. Чтобы добыть себе пропитание, хищники используют самые разные тактики. Добыча в свою очередь фатализмом не отличается и хочет жить, а потому также придумывает новые тактики избегания нежелательных встреч. Во многом тактики и тех, и других часто схожи: маскировка, яды, группирование особей, эхолокация и т.д. Это коэволюция и есть хрупким балансом между жизнью и смертью, между съесть и быть съеденным, между процветанием вида и его гибельную. Взглянув на обитателей морских глубин, можно выделить несколько существ, чьи тактики избегания неминуемой смерти, мягко говоря, странные. К примеру, морской огурец может вывернуть наизнанку свои собственные кишки, дабы отпугнуть неприятеля. А некоторые рыбы-попугаи заворачиваются перед сном в кокон из слизи, скрывающий их запах от хищников. А что морские ежи? Их внешний вид наталкивает на мысль, что иголки и есть их основной метод защиты. Это так, но он не единственный. Ученые из Барселонского университета (Испания) установили, что морские ежи, несмотря на свой малоподвижный образ жизни, способны «убегать» от опасности по вполне прямой траектории. Как существо без глаз, носа и ушей понимает, куда бежать, насколько траектории побега логичны, а не случайны, и помогает ли побег выжить? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Что потенциальной добыче нужно, чтобы убежать от хищника? В первую очередь, конечно, знать, что этот хищник в опасной близости от нее. Разные виды существ используют разные механизмы (и их комбинации) обнаружения хищников: слух, обоняние, зрение, тактильное ощущение и т.д. Но самым надежным, по мнению многих ученых, является химический метод обнаружения.
Способность определить по химическому следу факт присутствия неподалеку хищника или же факт того, что он был в этой области, позволяет не просто убежать от опасности, а избежать столкновения с ней. Подобный механизм обнаружения присущ как сухопутным, так и морским обитателям.
Существует два основных варианта развития событий. В первом случае химический сигнал хищника обнаруживается заранее, а потому у добычи есть время скрыться из виду, спрятаться где-то, т.е. найти убежище. Это одна из самых распространенных ответных реакций. Во втором случае времени искать укромный уголок уже нет, а потому наступает время банального (но единственно верного) варианта спасения — побега.
При наличии вариантов действий перед добычей стоит сложный выбор, так как нужно взвесить затраты энергии на побег или поиск укрытия. Следовательно, противодействие хищникам является типичным примером того, как животному необходимо быстро обработать информацию из окружающей среды, чтобы запустить соответствующую поведенческую реакцию (самую выгодную), которая ограничивается биомеханикой, возможностями обработки информации и состоянием тела животного (наличие болезней или травм, которые могут его замедлить).
Классическая формулировка взаимодействий хищник-добыча (например, модель Лотки-Вольтерра) предполагает, что хищники и жертва взаимодействуют случайным образом, подобно молекулам в идеальном газе, претерпевающем броуновское движение. По такой логике вероятность их встречи зависит от их концентраций (плотностей). Но в такой модели ни хищники, ни добыча не получают какой-либо внешней информации, а их взаимодействие происходит только в момент пересечения их траекторий движения.
Ученые отмечают, что многие исследования сосредоточены на том, как хищники находят и ловят добычу, но не затрагивают модели передвижения на индивидуальном уровне. В одном из немногих примеров, где этот аспект был включен, показано, что режим охоты хищника (мобильность или ожидание на месте) контролировал успех антихищнического поведения жертвы, что влияло на стабильность сообщества. Другими словами, поведение хищника влияет на успешность добычи в побеге от оного. Однако в таких моделях внимание уделяется только хищникам, предполагая при этом, что движения добычи случайны.
В современных трудах все больше внимания уделяется начальным этапам побега добычи от хищника. В расчетах используются направление, скорость побега и ускорение. Теоретические модели показывают, что на основе относительных скоростей хищника и жертвы можно спрогнозировать единую оптимальную траекторию побега.
Однако эти предсказания не учитывают важный аспект побега — его непредсказуемость. С эволюционной точки зрения, если добыча желает избежать хищника, ее тактика не должна быть однотипной, иначе второй быстро освоиться и учтет ее в будущей охоте.
Конечно, первые миллисекунды атаки хищника являются часто решающими как для него, так и для судьбы добычи. Нередко в этот короткий период времени наблюдается классическое поведение добычи — быстрое отступление. Однако следом может идти фаза так называемого «изменчивого поведения» с непредсказуемыми траекториями побега.
Потому ученые считают, что для лучшего понимания побега от хищников необходимо учитывать не только первые этапы этого побега, но и всю его траекторию. Кроме того, чтобы объединить поведение с динамикой популяции, необходимо охарактеризовать не только поведение «поиск добычи» у хищника, но и понять, что траектории побега жертвы могут отклоняться от моделей идеального газа, предложенных в прошлом.
Для этого ученые выбрали весьма необычного испытуемого — морского ежа (Echinoidea). Данный класс иглокожих насчитывает порядка 950 видов, которых можно встретить во всех океанах и в прибрежной, и в глубоководной зоне (5000 м). По примерным оценкам эти необычайные существа обитают на Земле уже порядка 450 миллионов лет.
Тело ежа имеет сферическую (или почти сферическую форму) диаметром от 2 до 30 см и покрыто рядом известковых пластин, соединенных друг с другом, формируя панцирь. В зависимости от формы тела ежей подразделяют на правильных и неправильных. У первых форма тела это сфера с радиальной пятилучевой симметрией. У неправильных — сфера сплющена, а потому можно различить передний и задний концы тела ежа. Образ жизни у морских ежей малоподвижный, а питаются они растительностью или сидячими (подвижными) видами животных.
Отличительной чертой морских ежей, как это видно из названия, являются иголки, служащие для передвижения, защиты и добычи пищи. Иглы могут быть разной формы и габаритов, в зависимости от функции. Кроме иглы есть и амбулакральная система, необходимая для движения, дыхания, выделения и осязания. Она состоит из околоротового кольца и пяти радиальных амбулакральных каналов. С внешней средой система контактирует посредством мадрепоровой пластинки. Также на теле некоторых ежей встречаются маленькие щипчики (педицеллярии) из 2 или более створок, необходимые для защиты и для очистки тела от мусора. Иглы некоторых видов оснащены ядом.
Самым же удивительным аспектом биологии морских ежей является их сенсорная система. Одни виды способны различать светлые и темные объекты, другие демонстрируют химиосенсорные механизмы обнаружения хищников и даже раненых сородичей.
Изучение ответов на стимулы, полученные одним из вышеперечисленных путей, в моделях взаимоотношений хищникдобыча часто ограничены сравнение угла движения до и после обнаружения хищника и пройденного расстояния. При этом никто не уделял внимание изменениям в траектории побега. Авторы рассматриваемого нами сегодня труда решили исправить этот недочет.
Подготовка к опытам

Paracentrotus lividus
В своем исследовании они изучили в лабораторных условиях долгосрочные (от минут до часов) углы курса, траектории, диффузионные свойства и предсказуемость движения морского ежа вида Paracentrotus lividus. Также была проведена оценка роли химических сигналов хищника в формировании поведения морского ежа.
В экспериментальных наблюдениях использовались морские ежи Paracentrotus lividus (диаметр тела 3.6 ± 0.05 см без игл) и хищные брюхоногие моллюски Hexaplex trunculus (длиной около 8 см).

Hexaplex trunculus
Камера для наблюдений состояла из металлического круглого резервуара (диаметром 3 м), облицованного пластиком. На высоте трех метров над ареной была установлена камера Nikon D80 с объективом 17 мм, позволяющая полностью видеть всю экспериментальную камеру.
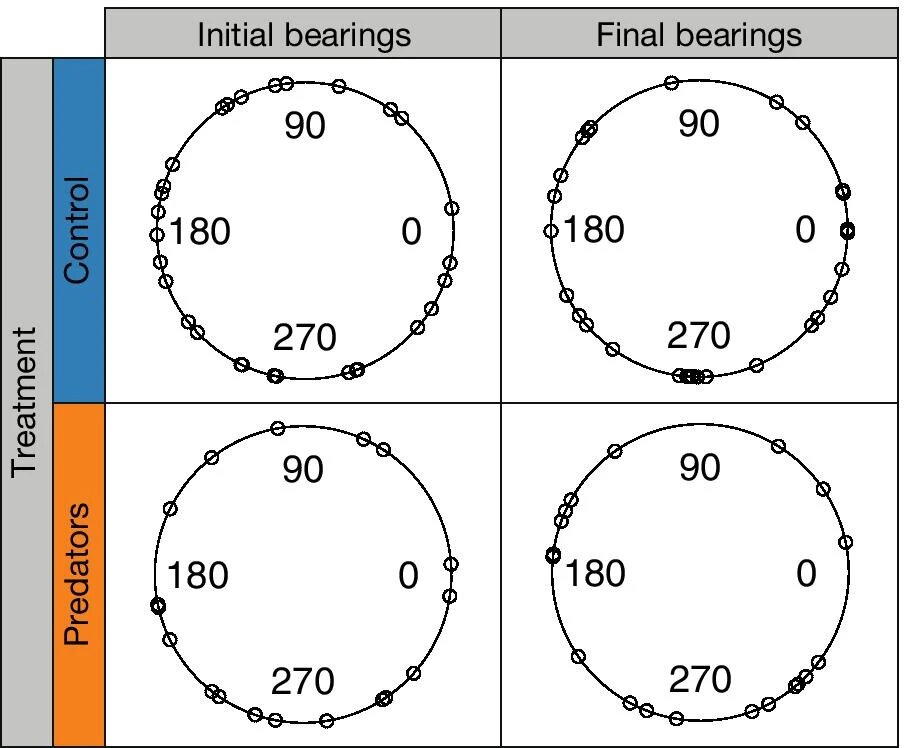
Изображение №1: распределение начальных и конечных углов направления морских ежей в контрольной и тестовой группах.
Каждый день испытаний камеру заполняли либо фильтрованной морской водой той же температуры и солености, что и в аквариумах, где временно обитали испытуемые, либо морской водой, содержащей химический сигнал моллюска Hexaplex trunculus. Морская вода с химическим сигналом была получена из аквариума с моллюсками, где 6 особей пребывали в течение ночи.
Каждую особь морского ежа тестировали всего раз. В начале каждого испытания морского ежа помещали в центр камеры. Опыт считался завершенным, когда еж достигал расстояния в 10 см от края камеры. В чистую морскую воду (контрольная группа) поочередно помещалось 29 особей, а в воду с химическим сигналом (тестовая группа) — 21 особь.
Результаты экспериментов
Траектории морских ежей, подвергшихся воздействию химических сигналов хищников, в среднем были более прямыми (на 28%) и более быстрыми (на 34%), чем у контрольной группы (2a, 2b).
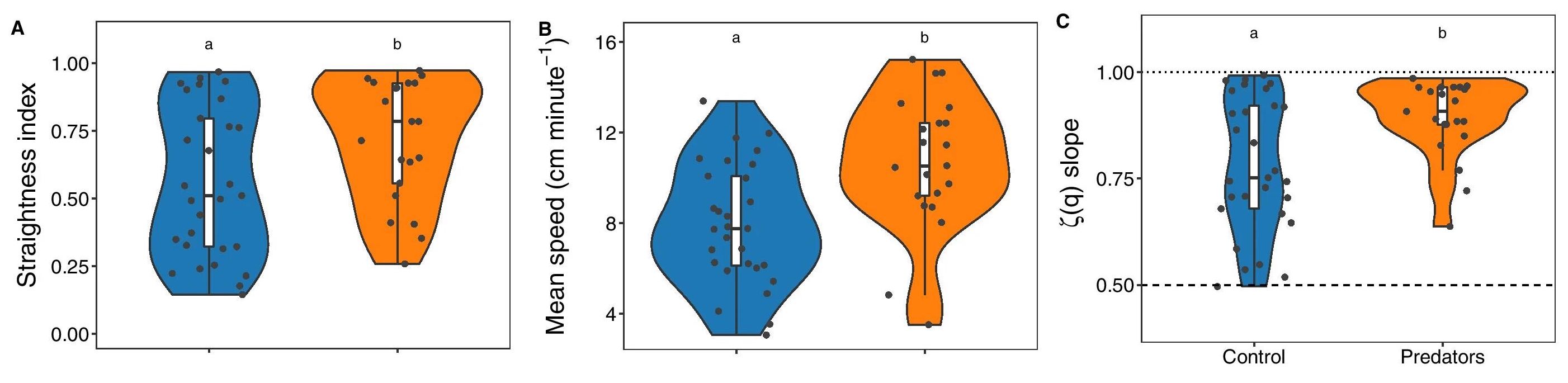
Изображение №2
Показатели масштабирования моментов q-го порядка (ζ(q)) позволили оценить диффузионные свойства траекторий отдельных морских ежей (2c, 3b и 3d).
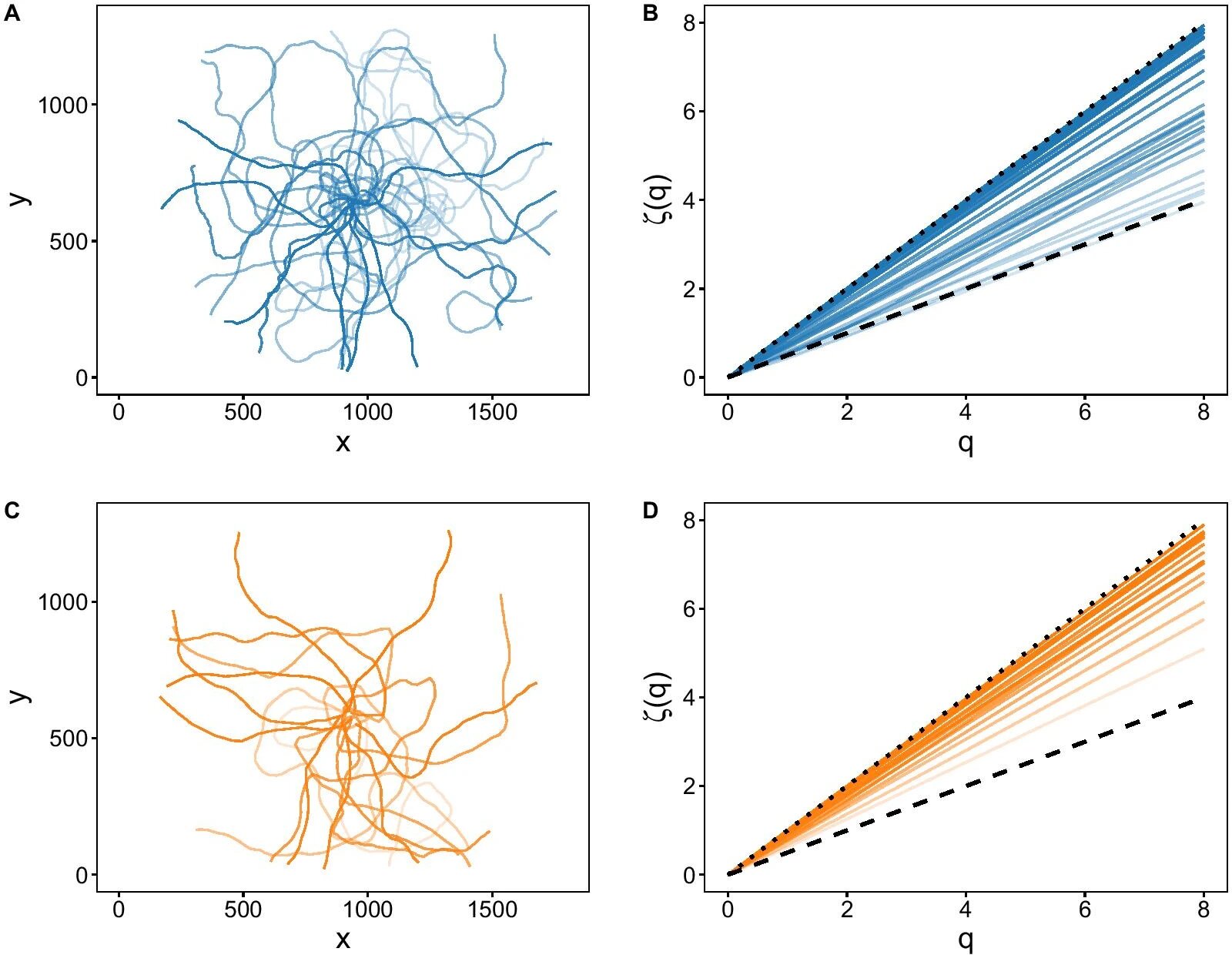
Изображение №3
На 2c, 3b и на 3d самые крутые склоны (самые высокие коэффициенты наклона) соответствуют баллистическим траекториям (баллистическое движение происходит при наклоне ζ(q) = 1; точечная пунктирная линия), а самые пологие склоны соответствуют броуновскому движению (при наклоне ζ(q) = 0.5, штриховая пунктирная линия). А наклоны между броуновской и баллистической областями соответствуют супердиффузионным траекториям.
Диффузионные свойства траекторий морских ежей из контрольной группы сильно варьировались. Тогда как в присутствии сигналов хищников диапазон поведения морского ежа был ограничен (сравнение фигур на 2c и сравнение графиков 3b с 3d).
Проще говоря, в контрольной группе движение ежей было хаотичным (броуновским), но иногда встречались и прямые траектории. Но вот в тестовой группе подавляющее большинство траекторий были именно прямыми.
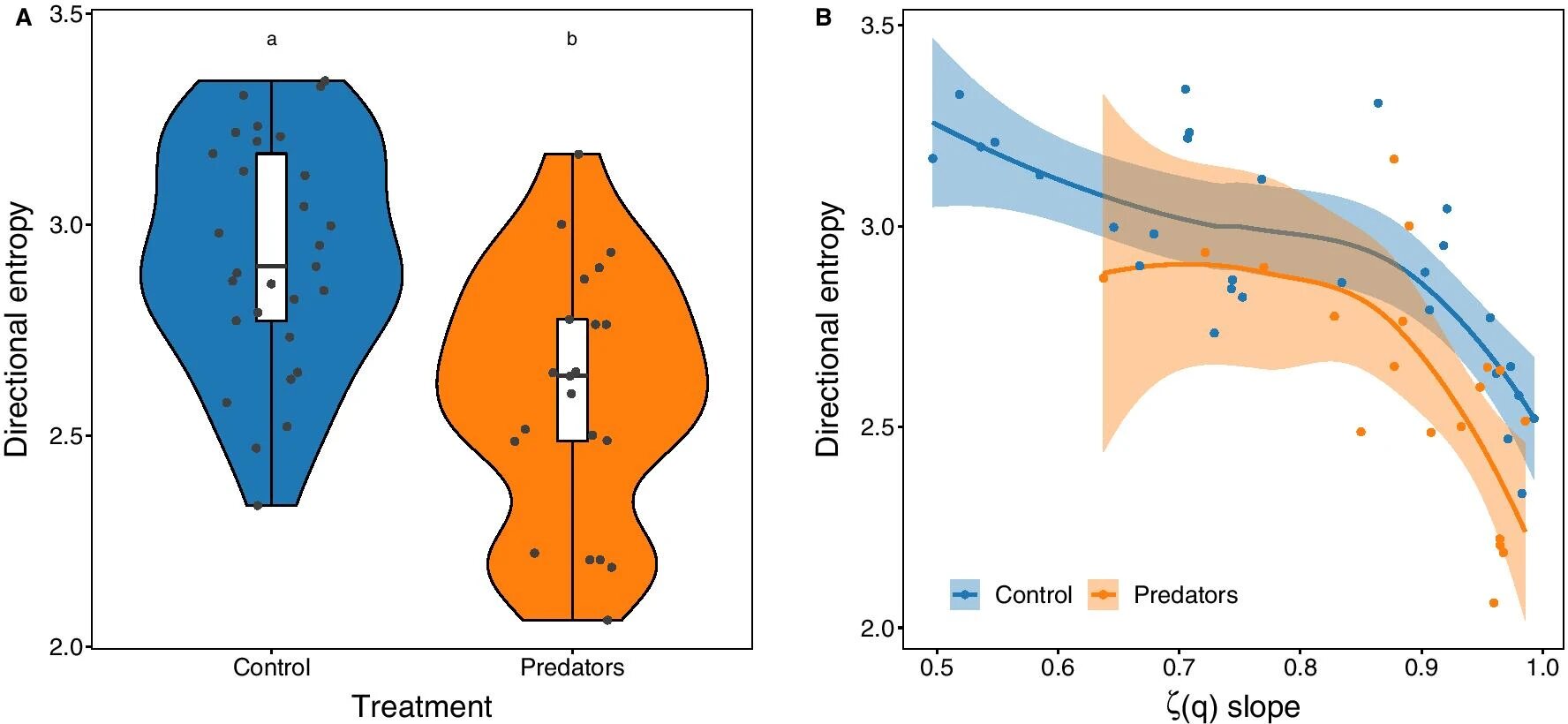
Изображение №4
Помимо вышеописанного отличия поведение тестовой группы можно было намного легче и точнее предсказать. Энтропия направления индивидуальных траекторий морских ежей от воздействия сигналов хищников снизилась в среднем на 13% по сравнению с контрольной группой (4а). Энтропия направления морских ежей уменьшалась с увеличением крутизны функции ζ(q). Это означает, что траектория переходила от броуновского движения к баллистическому, а предсказуемость такого перехода и дальнейшего развития траектории возрастала (4b).
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
Вышеописанные результаты показывают, что у морских ежей действительно есть сложные паттерны движений. Это удивительно, если учесть ограниченность получаемой ежами информации, ввиду отсутствия у них полноценных органов чувств. Опыты показали, что страх перед хищником является крайне важной движущей силой изменения траектории морского ежа. Следовательно, у этих существ есть несколько моделей поведения, соответствующих различным событиям в их жизни: траектория движения, оптимизированная для максимально эффективного поиска и добычи пищи, и специфическая траектория на случай контакта с хищником.
По словам ученых, их находка является веским доказательством того, что существующие модели поведения хищник-добыча, которые предполагают хаотичное и непредсказуемое движение обоих, являются неверными или как минимум неточными.
В будущем ученые намерены продолжить работать над своим исследованием. В частности они хотят провести моделирование, в котором будет учтена наблюдаемая в данном труде сложная модель движения морских ежей. Это поможет сделать модель хищник-добыча гораздо точнее, что в свою очередь поможет лучше понимать и предсказывать динамику в популяциях обоих.
Также остается важный вопрос — если морской еж, будучи далеким от статуса самого развитого организма на планете, демонстрирует сложную динамику поведения в разных условиях, то что было не учтено в моделировании поведения более развитых животных.
Стоит также отметить, что данный труд косвенно показал, насколько порой бывают важны детали, и исключение того, что кажется неважным, может привести к совершенно неточным результатам.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


