
На необъятных просторах нашей галактики сокрыто множество секретов, которые так усердно пытаются найти и разгадать ученые со всего мира. Однако не обязательно чему-то быть большим, чтобы быть загадочным. Ярким тому доказательством является мир, лежащий на клеточном уровне. Множество самых разных по форме, строению, функционалу и назначению клеток совместно выполняют общую задачу — поддержание жизни организма. Если утрировать, то у клеток, как у людей, есть профессии: почтальоны, передающие информацию между клетками и тканями; пограничники, выявляющие и борющиеся с инфекциями; архивариусы, собирающие и хранящие информацию и т.д. В этом невероятном спектре специальностей есть весьма необычная, по крайней мере для нас, профессия — биологический мотор, которые генерирует механическое усилие, необходимое для движения клеток.
Эти клетки особенно интересны в контексте нанотехнологий. Ранее в реализации работоспособного нано-устройства на базе биологических моторов была проблема — моторы должны быть интегрированы в более крупные системы, чтобы их механические движения могли быть эффективно связаны с другими молекулярными единицами. Ученым из Мюнхенского университета (Германия) удалось приблизиться к реализации этой концепции. Какие именно клетки и молекулярные единицы были использованы в создании модели, как контролировалась их работа, какова была задача работающей системы и какие результаты она показала? Ответы на эти вопросы мы найдем в докладе исследовательской группы. Поехали.
Основа исследования
Как уже было сказано выше, биологический мотор это клетка, которая генерирует механическое усилие, необходимое для реализации движения клеток, а также внутриклеточного транспорта. К таким моторам относят моторные белки и белковые комплексы.
Чтобы осуществлять свою нестандартную деятельность моторным белкам, как и любой машине, нужно топливо. Его роль исполняет аденозинтрифосфорная кислота (АТФ, C10H16N5O13P3). АТФ является универсальным источником энергии для всех биологических процессов внутри живых систем.
Двигательные белки функционируют за счет гидролиза* АТФ, что позволяет белкам преобразовывать химическую энергию в механическую работу.
Гидролиз* — реакция обменного разложения между растворенным веществом и растворителем, при которой происходит разложение вещества (в данном случае АТФ) и воды с образованием новых соединений.
В частности к моторным белкам относят миозины, кинезины и динеины. Миозины составляют порядка 40-60% от всех мышечных белков, участвуя в процессе мышечного сокращения.
Кинезины, ставшие научным мемом несколько лет назад, перемещаются по микротрубочкам (белковые внутриклеточные структуры), участвуя в процессах митоза, мейоза и везикулярного транспорта.
Видео-демонстрация того, как кинезин движется по микротрубочке, преобразованная в мем. (оригинал взят из доклада The Inner Life of a Cell; музыка, наложенная на видео — «Stayin’ Alive», Bee Gees, 1977 год).
Динеины, как и кинезины, также перемещаются по микротрубочкам цитоскелета, участвуя в процессе переноса груза (везикулы, митохондрии и т.д.).
Интерес к биомоторам со стороны нанотехнологов обусловлен несколькими важными факторами: наноразмерность, биосовместимость и возможность использовать генную инженерию для создания биомоторов с заданными функциями.
На данный момент существует ряд разработок, основанных на принципах действия биологических моторов. Однако информация касательно того, какой реальный объем работы может выполнять один молекулярный мотор, пока недостаточно для полноценной реализации. Еще одной головоломкой для ученых является вопрос о том, как интегрировать молекулярный двигательный блок в более крупную структуру так, чтобы его направленное движение эффективно трансформировалось в повышение потенциальной энергии в удаленных точках структуры.
Очевидно, что преград много, но это ученых еще никогда не останавливало. В рассматриваемом нами сегодня исследовании ученые описывают молекулярную установку 1, которая позволяет передавать потенциальную энергию от моторного блока к удаленному блоку-приемнику, за счет чего ускоряется движение последнего (изображение №1).
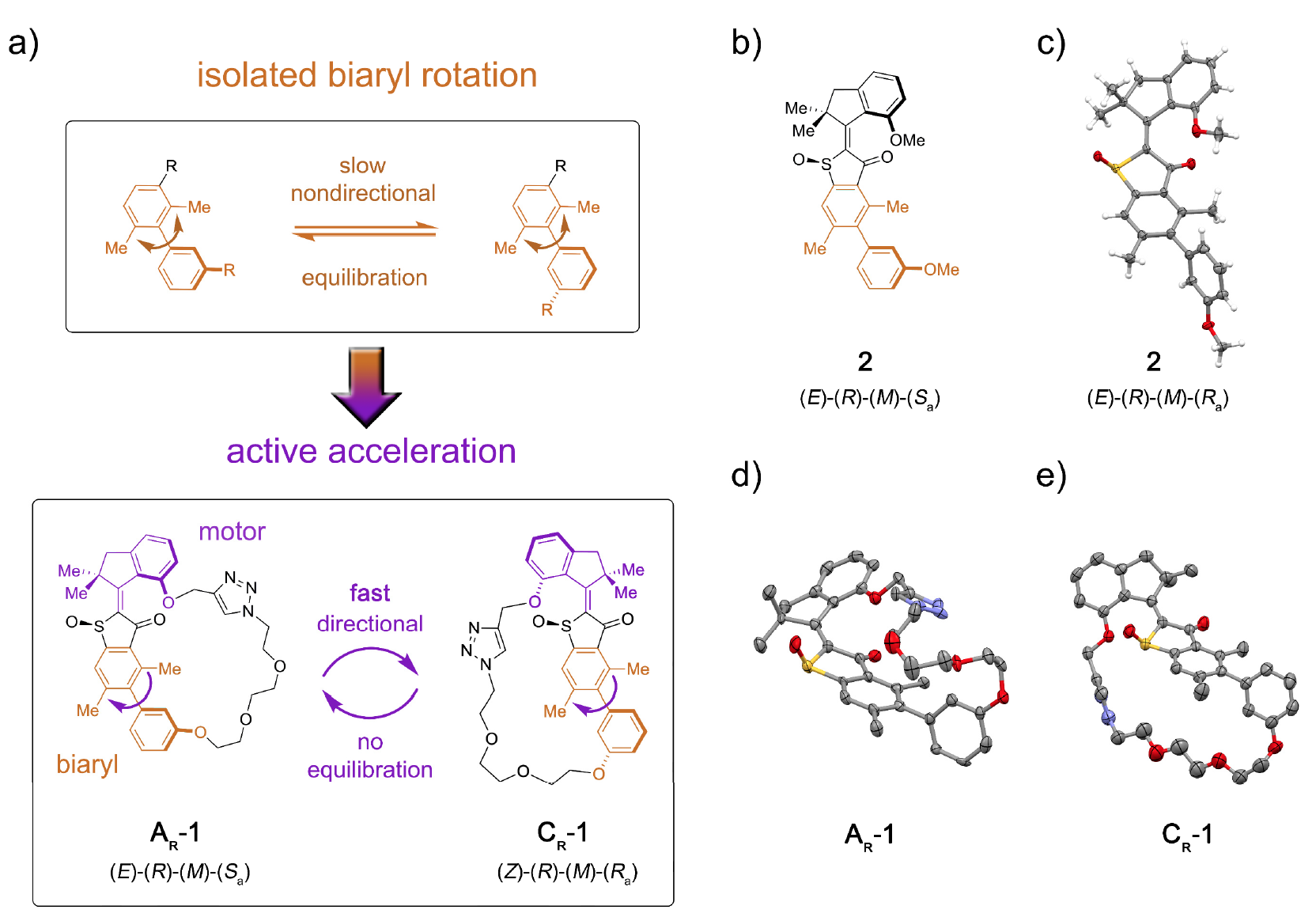
Изображение №1
Роль блока-приемника исполнил аксиально-хиральный* биарил*, который в его нететеризованной форме (модельная система 2) подвергается медленному и ненаправленному вращению атропоизомеризации в направлении равновесия энантиомеров 1: 1 (1а).
Аксиальная хиральность* возникает в результате неплоского расположения заместителей относительно некоторой оси — оси хиральности.
Биарил* — Любое соединение, содержащее субструктуру, которая представляет собой совокупность двух ароматических соединений или арильных групп, если они соединены одинарной связью.
Сопряжение молекулярного моторного блока позволяет однонаправленно отвести эту атропоизомеризацию от равновесия. В результате атропоизомеризация биарила больше не является пассивной (следуя работе мотора), а представляет собой энергетическое препятствие, против которого мотор должен активно работать.
В машине 1 увеличение скорости атропоизомеризации биарила на несколько порядков достигается действием мотора, подавляющего внутренние барьеры для изолированного вращения биарила (модельная система 2 на 1b и 1с).
В молекулярную установку 1 входит молекулярная моторная единица, основанная на HTI, относящемуся к классу индигоидных хромофоров. Данный тип единиц крайне чувствителен к свету, т.е. благодаря свету ею можно управлять. Моторный блок ковалентно соединен с удаленной биарильной осью, которая не движется вместе с вращением мотора.
Одним из отличий данной установки от более ранних ее вариантов является наличие дополнительных состояний (шагов): ранний вариант имел 4 состояния и регулировался только шагами вращения мотора; новый вариант — шестиступенчатая система (изображение №2).
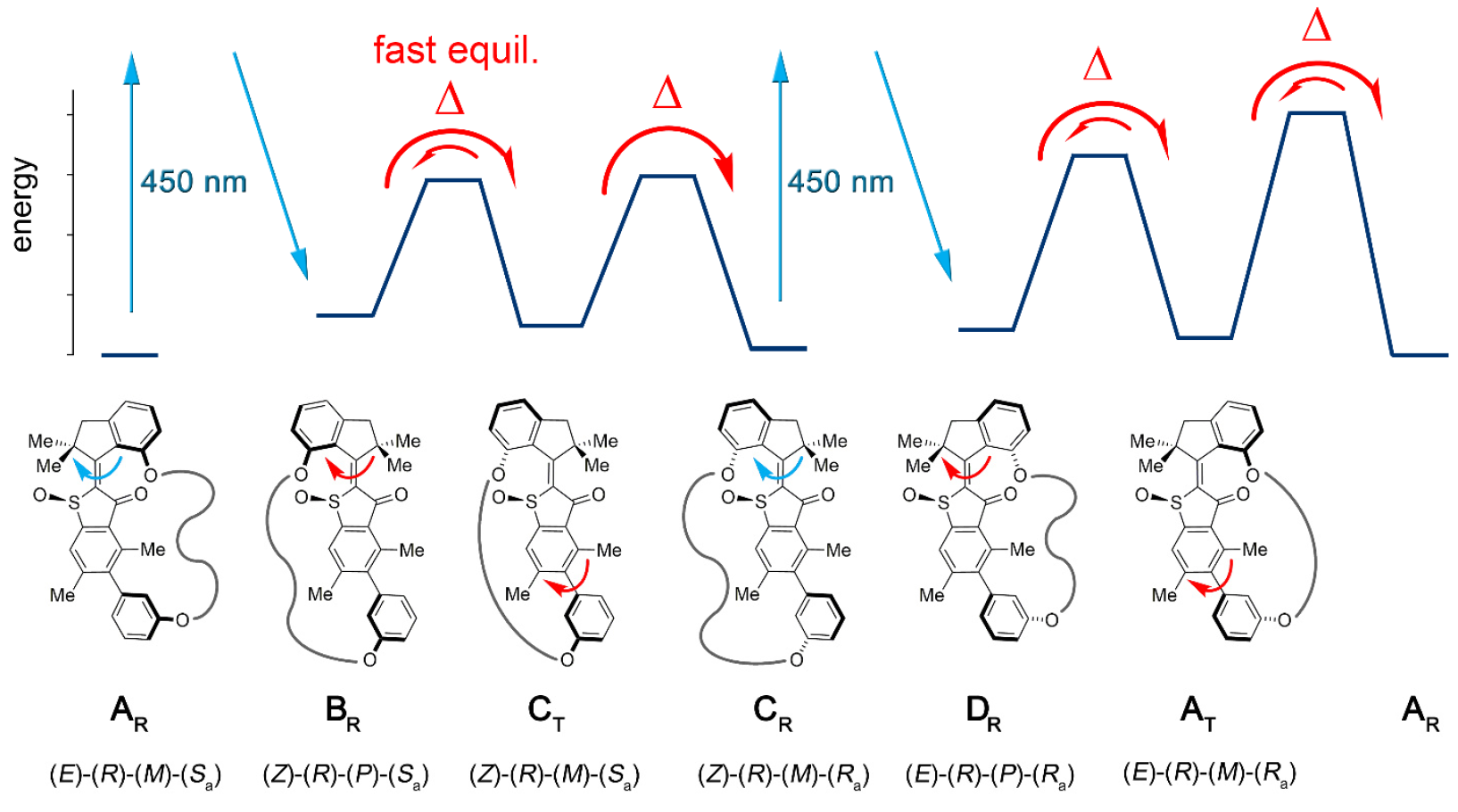
Изображение №2
Шесть изомерных состояний названы следующим образом: AT для напряженного состояния A; AR для расслабленного состояния A, а также BR, CT, CR и DR (промежуточные состояния системы).
Пять из этих шагов можно наблюдать экспериментально, что подтверждает наличие шестого шага и, следовательно, подтверждает полную направленность для вращения на 360 градусов связанной системы блок-мотора и блок-получатель.
Моторная система 1 была синтезирована конвергентным методом за счет бромированного HTI-предшественника, к которому посредством катализируемой медью клик-реакции присоединена ковалентная связь, содержащая функцию сложного боронового эфира.
Впоследствии реакция Сузуки* дает макроциклизацию*, за которой следует окисление с получением окончательной структуры 1.
Реакция Сузуки* — органическая реакция арил- и винилборных кислот с арил- или винилгалогенидами, катализируемая комплексами Pd(0).
Макроциклизация* — реакция циклизации, которая приводит к формированию макроцикла, т.е. химического соединения, в котором имеется 9 или более связанных атомов, образующих кольцо.
Для двух наиболее стабильных состояний AR и CR системы 1 и наиболее стабильного состояния системы 2 были получены кристаллы, подходящие для структурного анализа (®-конфигурированные изомеры рацемического* AR на 1d; CR на 1е).
Рацемат* — эквимолярная смесь двух стереоизомеров, представляющих собой зеркальные отражения друг друга. Рацемическое соединение состоит из кристаллов, в каждом из которых есть молекулы обоих энантиомеров, а их соотношение равно 1:1.
Поскольку AR/AT и CR могут быть разделены с помощью ВЭЖХ (высокоэффективная жидкостная хроматография), их поведение в условиях нагревания и облучения при различных температурах может быть изучено независимо. При нагревании раствора рацемического CR в (CDCl2)2 до 80 °C — 140 °C более стабильный AR образовывался в 93% случаев. Это устанавливает разность свободной энтальпии ∆G = 1.8-1.9 ккал/моль в этом интервале температур между этими двумя состояниями. Соответствующий кинетический анализ выявил высокоэнергетический барьер в 28.2 ккал/моль при 80 °С, сопровождающий термическую изомеризацию двойной связи Z/E.
При охлаждении чистого CR до -105 °C в CD2Cl2/CS2 (соотношение 4/1) и облучении его светом 450 нм возникает новый набор сигналов, который отличается от известного набора сигналов AR и AT (3а).

Изображение №3
Сигналы изомера DR затухают до 75% в течение 28 минут при температуре — 80 °C и полной темноте, а вот сигналы изомера AT, напротив, растут (3е).
Тепловое равновесие между DR (оставшиеся 12%) и AT (88%) наблюдается при -60 °C, что выражается в ∆G = 0.84 ккал/моль между двумя состояниями. Кинетический анализ термического распада выявил сопутствующую свободную энтальпию активации ∆‡G = 13.9 ккал/моль при -80 °C для этого процесса. При температуре от — 40 до 0 °C наблюдается ослабление сигналов AT и сопутствующий рост известных сигналов AR до момента достижения равновесия между ними. Кинетический анализ этого процесса определил ∆‡G, равную от 18.4 до 19.3 ккал/моль при температуре от — 40…0 °C.
Таким образом, было установлено, что облучение CR приводит к первому продукту фотоизомеризации DR, который термически превращается в изомер AT посредством инверсии единственной спирали в моторном блоке. При охлаждении равновесного раствора AR/AT в CD2Cl2/CS2 до — 105 °C и при его облучении светом 450 нм наблюдалась фотореакция AR, что объясняется его распространенностью. Также появляется новый набор сигналов, отличающихся от уже известных сигналов CR (3f). Эти новые сигналы, будучи продуктом фотореакции AR, подтверждают ожидаемую направленность фотоизомеризации от AR к BR, которая затем еще больше стабилизируется, подвергаясь быстрой спиральной инверсии к CT даже при низкой температуре.
При температуре — 80 °C и при полной темноте сигналы BR/CT практически полностью исчезают, и лишь сигналы CR продолжают усиливаться и дальше (3е). Это дает нижний предел для разницы энергий между CT и CR, равное 0.98 ккал/моль.
Кинетический анализ определил ∆‡G = 13.4-13.5 ккал/моль, необходимую для превращения СТ в CR. Видно, что энергия активации атропоизомеризации СТ в CR снижается по сравнению с этим показателем для атропоизомеризации AT в AR (18.4 ккал/моль). Это может быть связано с большим напряжением (деформацией) в СТ по сравнению с АТ, что может быть связано с большим расстояниям между двумя фенольными атомами кислорода, служащими точками соединения линкерной цепи в структурах С, чего нет в структурах А. В кристаллическом состоянии CR расстояние между двумя атомами кислорода составляет 10.6, тогда как в AR — 7.6. Это дает линкерной цепи большую конформационную (пространственное положение атомов) свободу в А-структурах, нежели в С-структурах. Если же учесть свободную энтальпию активации для термического превращения CT в CR, то этот процесс превращения может быть полностью остановлен при -105 °C.
При температуре -105 °C можно проверить степень накопления СТ при сильном облучении светом AR. Подобное возможно, если разница энергий между BR и СТ превышает 2.0 ккал/моль и если СТ структура не участвует в каких-либо процессах фотохимии сама по себе. Однако накопление СТ при облучении AR при -105 °C невозможно, так как имеется быстрое тепловое уравновешивание между СТ и BR.

Таблица №1
Совокупность вышеописанных наблюдений и данных позволяет создать механическую картину 6-ступенчатой последовательности однонаправленного вращения в модели 1 (изображение №2 и №4).

Изображение №4
Эксперименты с разными температурными режимами позволили определить температурную зависимость ∆G между CR и AR/АТ, DR и AT, AT и AR, а также температурную зависимость ∆‡G для BR/CT и CR, DR и AT, AT и AR (таблица №1).
В отличие от обычного 4-ступенчатого механизма на базе HTI мотора полное вращение на 360 градусов включает в себя два дополнительных шага, которые представляют собой повышение потенциальной энергии, передаваемое от мотора к биарильному блоку. Во время этих двух шагов формируется энергетическая связь между мотором и биарильным блоком (4b), т.е. последний более не является энергетически вырожденным.

Изображение №5
Вместо этого энергия любого из атропоизомеров растет за счет «храповых» шагов мотора во время одного полного вращения (5с и 5d). Это энергетическое изменение обеспечивает до 90% взаимопревращения атропоизомера на следующих термически активируемых стадиях и, следовательно, почти полное вращение биарильного блока в течение одного рабочего цикла механизма.
Это наблюдение может говорить о том, что биарильный блок претерпел повышение потенциальной энергии, что является результатом работы моторного блока. Стоит отметить, что изначальное повышение энергии значительно больше, чем то, что доходит до биарильного блока. Так, лишь 72% от нее передается от мотора к биарильному блоку.
Вышеописанные наблюдения говорят о том, что длина соединения между мотором и биарильным блоком является важным аспектом в процессе изменения степени передачи энергии. Следовательно, более короткое соединение может позволить передать больше потенциальной энергии от BR и DR к АТ и СТ.
Для более подробного ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В данном труде ученые описали принцип работы своего изобретения, основанного на взаимодействии биологических моторов и молекулярных элементов. Разработанная установка позволяет передавать потенциальную энергию от мотора к определенному элементу (в данном случае это биарильный блок). В результате активности мотора термическая атропоизомеризация биарила вынуждена протекать однонаправленно и с большей скоростью. Передача энергии происходит во время термически активируемых храповиков (шагов) в механизме вращения. Также было установлено, что порядка 72% от первоначальной энергии мотора передается на заданный блок.
Данное изобретение является важным шагом на пути создания полноценных биологических нанометровых устройств. Использование био-моторов в проектировании таких механизмов обусловлено их биосовместимостью, легкостью в изменении функционала за счет генной инженерии, а также их наноразмерностью от природы. Искусственные моторы на данный момент не могут похвастаться вышеперечисленной комбинацией преимуществ.
Исследователи говорят, что следующим шагом в их работе станет демонстрация того, что их установка способна выполнять не только абстрактные действия, но и быть полезной. Другими словами, их будущее творение будет более усовершенствовано для выполнения полезных функций на молекулярном уровне.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята. 🙂
Немного рекламы 🙂
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


