 Уже давно бьют тревогу по поводу возникновения резистентных к антибиотикам штаммов бактерий, хотя некоторые серьёзные заболевания являются вирусными и также требуют скорейших мер по взятию их под контроль.
Уже давно бьют тревогу по поводу возникновения резистентных к антибиотикам штаммов бактерий, хотя некоторые серьёзные заболевания являются вирусными и также требуют скорейших мер по взятию их под контроль.
К таким заболеваниям относятся, конечно, СПИД, вызываемый вирусами иммунодефицита человека, гепатит С, который, если его не лечить, во многих случаях может приводить к злокачественным новообразованиям печени и циррозу, герпес, грипп типа А, бешенство, геморрагическая лихорадка Эбола и некоторые другие.
Принципиальное отличие жизненнного цикла вируса от жизненного цикла бактерии заключается в том, что вирус — это облигатный внутриклеточный паразит. Фактически вирус вне клетки мёртв, так как не может ни размножаться, ни самостоятельно перемещаться, ни демонстировать каких-либо иных признаков жизни. Вся его жизнь, которая заключается исключительно в воспроизведении себе подобных, протекает только при попадании в вожделенное начало всех начал — клетку.
Безусловно, именно этот факт и определяет мишени фармакологического воздействия при вирусных заболеваниях.
В фармакологии инфекционных и паразитарных заболеваний (отчасти это справедливо и для случая злокачественных новообразований), существует 2 принципиально различных типа лекарственного воздействия. Одно из них направлено на модификацию макроорганизма-хозяина, то есть человека, с целью выработки устойчивости к инфекционному агенту или усиления защитных свойств организма. Классический пример — вакцинация. Второй тип лекарственного воздействия направлен на модификацию (ослабление, уничтожение) паразита, то есть патогенного агента. Классический пример этого типа – применение антибиотиков.
Меня, как химика, прежде всего интересует использование при вирусных заболеваниях так называемых малых молекул, которым и будет посвящена данная статья.
Так что же дают медицинскому химику, который занимается конструированием лекарственных средств, знания в области вирусологии? Прежде всего, они позволяют определить точки приложения лекарственной молекулы. Вот и пройдёмся по ним.
Представьте, что вы — попавший в организм вирус. Перед вами — пустыня, почти голая, почти безжизненная клетка с мембраной. Что будете делать? Понятно, что в клетку как-то надо проникнуть. Но, сори, просто так вам туда войти никто не даст. Для диффузии, хоть простой, хоть облегчённой, вы слишком велики. Никаких специальных каналов и белковых-переносчиков вам матушка эволюция не оставила. А вы что хотели: чтоб как в сказке, да ещё с проницаемыми мембранами? Щаз.
Но, как говорится, и вы не лышком шиты. А вирусной оболочкой, или капсидом.
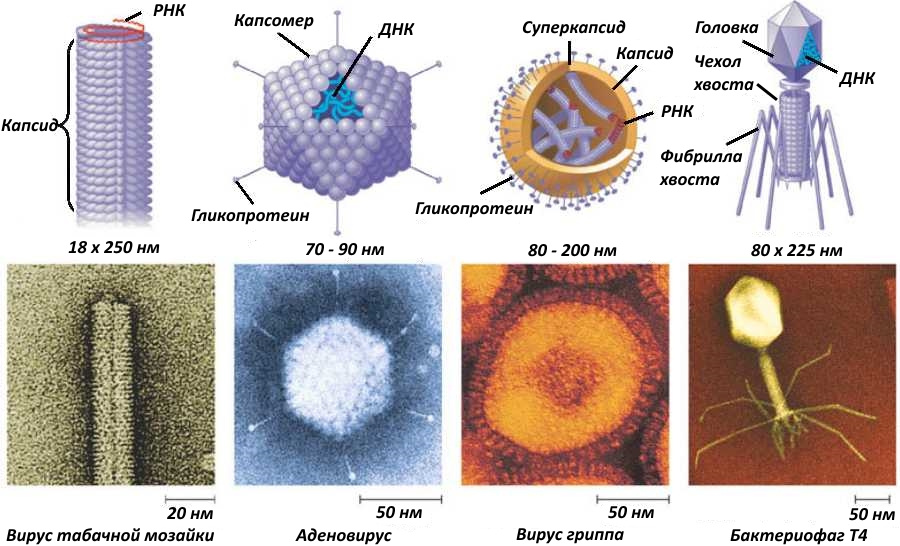
Хочу заметить, что капсид состоит из белков, но некоторые вирусы имеют и суперкапсид. Суперкапсид — это липидная оболочка, захваченная вирусом при выходе из клетки хозяина. Как она образуется у ВИЧ, можно посмотреть на рисунке ниже

В дальнейшем я буду писать «капсид», подразумевая в тех случаях, где речь идёт об оболочечных вирусах, что может быть и суперкапсид.
Капсид имеет на своей поверхности «отростки», чаще всего из гликопротеинов, которые связываются с каким-либо из многочисленых типов рецепторов, торчащих из клетки. Рецепторы эти, естественно, нужны вовсе не для проникновения вируса в клетку, а для получения сигналов из вне для нормальной жизнедеятельности.
К примеру, если вы — ВИЧ, то облюбуете себе СD4-рецептор, который встречается на поверхности клеток иммунной системы (лимфоцитов, моноцитов, макрофагов, клеток Лангерганса, дендритных клеток), а также клеток микроглии — по сути, компонента иммуной системы головного и спинного мозга.
Если вы домосед, не хотите переезжать с места на место в шастающих туда-сюда клетках иммунной системы, то всегда можно одеться в личину вируса гепатита В или С. В первом случае к вашим услугам предоставляется натрий-зависимый транспортёр желчных кислот, открывающий уютные комфортабельные аппартаменты гепацитов печени. В случае гепатита С, что, кстати, встречается в вирусном мире редко, для попадания в гостеприимный гепацит вирус использует несколько рецепторов.
Любителям альпийского высокогорья прохлады и свежего воздуха могу порекомендовать стать вирусом гриппа и облюбовать эпителий верхних дыхательных путей. В мир упоительного блаженства вам проложат путь остатки сиаловых кислот, которые прочно свяжутся с гемагглютинином на поверхности вашего капсида.
Короче, принцип понятен — выбираете курорт по своим предпочтениям, а дальше — дело техники.
Очевидно, что связывание частей капсида с рецепторами на поверхности клетки является слабым звеном вирусной агрессии и может быть использовано для фармакологической атаки. Следуя логике, можно предложить два подхода, каждый из которых имеет свои недостатки. Во-первых, можно заблокировать гликопротеин на поверхности вируса, а во-вторых — рецептор на поверхности клетки. Если пойти по второму пути, то мы рискуем нарушить нормальную жизнедеятельность: клетка перестанет получать сигналы из вне и загнётся. А если это клетка иммунной системы, то можно потерять и весь макроорганизм. Кажется, что проще и безопаснее заблокировать вирусный гликопротеин. И это действительно так. Однако вирусы (особенно РНК-содержащие) очень сильно изменчивы, то есть быстро мутируют, поэтому созданный препарат может вскоре перестать работать. Отсюда, кстати, следует вывод о том, что мишенью для препарата должны стать наименее вариабельные (изменчивые) части капсида.
Теперь немножко примеров из клинической практики.
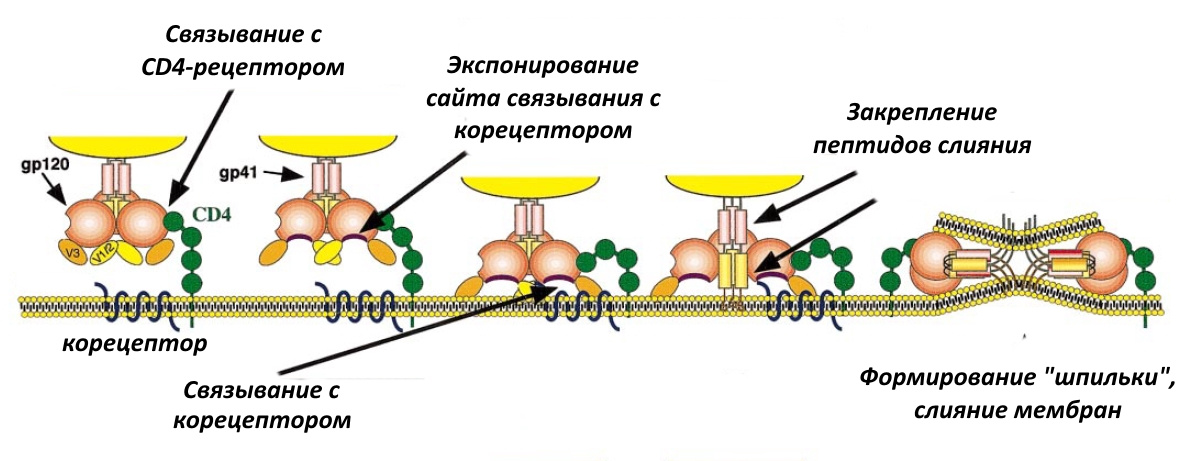
Существует 2 препарата, которые действуют на ВИЧ и препятствуют его попаданию в клетку. Первый, как это ни странно, использует химиотерапевтическую стратегию № 1, то есть модифицирует макроорганизм. Препарат называется маравирок и был разработан британским филиалом Pfizer в первой половине 2000-х. Механизм его действия такой. Чтобы проникнуть в клетку, ВИЧ использует не только CD4-рецептор, но и корецептор иммунных клеток, CCR5 (см. рис. выше). Маравирок изменяет пространственную структуру этого рецептора таким образом, что ВИЧ уже не в состоянии за него ухватиться и проникнуть в клетку.
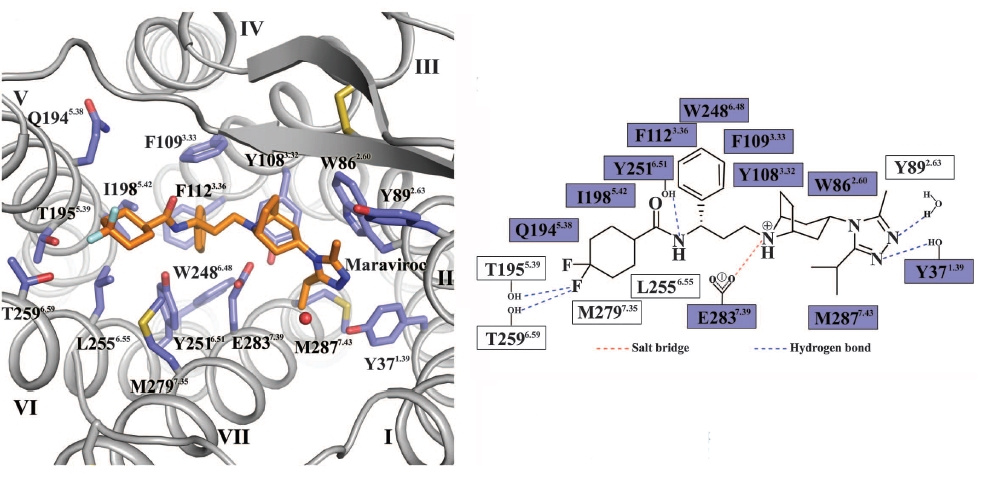
Обратите внимание, что молекула маравирока связывается с CCR5 как многочисленными водородными связями, так и электростатически.
Второй ингибитор слияния, как называют лекарства этого класса, энфувертид, не является классической малой молекулой, а представляет собой пептид — цепочку из из 36 аминокислот, первая из которых модифицирована остатком уксусной кислоты.

Она присоединяется к одному из гликопротеинов ВИЧ и блокирует его работу по внедрению частиц вируса в клетку на последнем этапе проникновения, когда происходит объединение суперкапсида (ВИЧ является оболочечным вирусом) и мембраны клетки. Весьма наглядно это показано в следующем ролике:
В разработке находятся ещё несколько лекарств из класса ингибиторов слияния, предназначенных для терапии ВИЧ и гепатитов. Посмотрим, насколько это направление окажется перспективным.
Продолжим.
Итак, на ресепшене вам дали ключи, и вы широко распахнули ворота клетки, призывно манящей своим содержимым. Что дальше?
Часть вирусов, и среди них ВИЧ, попадает после проникновения в клетку напрямую в цитоплазму, покинув остатки суперкапсида на поверхности цитоплазматической мембраны. Другие вирусы проделывают более сложный путь.
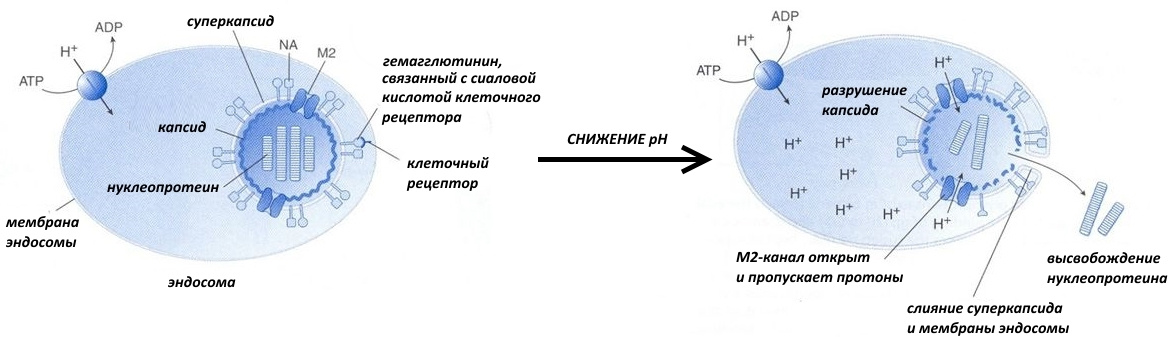
Так, связавшись гликопротеинами с клеточными рецепторами, вирус может попасть в клетку путём эндоцитоза, то есть вместе с выпячиваниями клеточной мембраны, формирующими пузырьки-эндосомы.
Со временем содержимое эндосомы закисляется, то есть его рН понижается. И тут вам за несколько сотен тысяч лет эволюции надо было успеть подсуетиться, а не щёлкать капсидом. К примеру, вирус гриппа создал специальный М2-белок.
Он представляет собой канал, по которому внутрь вирусной частицы устремляются протоны. В результате происходят следующие события:
— электростатическое взаимодействие между капсидом и нуклеопротеином (комплексом нуклеиновых кислот и белков) ослабляется, и они отсоединяются друг от друга;
— пептиды слияния встраиваются в стенку эндосомы, содержимое суперкапсида и цитоплазмы объединяется.
Таким вот образом в цитоплазму клетки попадает основная «рабочая» часть вируса, которая впоследствии захватывает весь свой гостиничный номер, — нуклеиновая кислота.
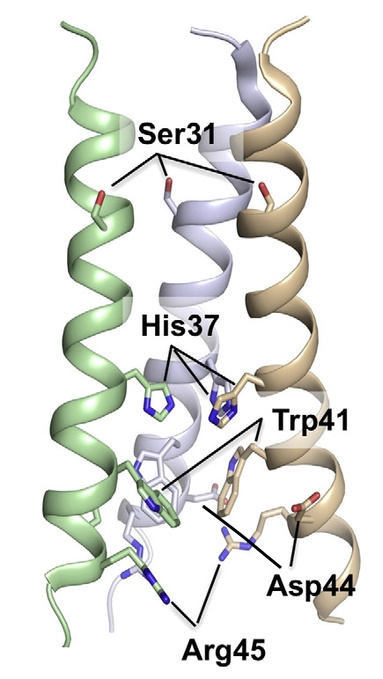
же есть. M2-белок представляет собой тетрамер, то есть состоит из 4 субъединиц и имеет 3 функциональных части. Трансмембранный сегмент образует проницаемый для протонов канал, а С- и N-концевые сегменты удерживают тетрамеры, чтобы те не «разъехались» при открывании канала.
Внутрь канала обращены остатки аминокислот гистидина (His37 или H37) и триптофана (Trp41 или W41), которые, как полагают, играют ключевую роль в обеспечении его протонной проводимости.
При физиологических значениях рН только 2 из 4 гистидиновых остатков протонированы, их имидазольные части находятся почти перпендикулярно α-спиралям тетрамеров.
Остатки триптофана также перпендикулярны α-спиралам. В таком положении им помогают удерживаться водородные связи пиррольных колец с остатками аспарагиновой кислоты.
При закислении содержимого эндосомы происходит дополнительное протонирование 1 или 2 остатков гистидина, силы электростатического отталкивания начинают превосходить
силы водородных связей, имидазольные фрагменты выходят из плоскости, перпендикулярной каналу, и открывают его для протонов.

Далее происходит поворот остатков триптофана в сторону протонированного имидазола по причине образования π-комплекса с его бензольной частью. Таким образом, триптофановые остатки также выходят из плоскости, их водородная связсь с остатками аспарагиновой кислоты разрывается, в результате чего протон проходит по образовавшемуся каналу далее вглубь вириона. Вуаля!

Итак, очевидно, что мы снова имеем болевую точку — раздевание капсида вируса гриппа при закислении эндосом. И этим грех не воспользоваться.
Заблокировать М2-белковый протонный канал можно с помощью знакомого вам римантадина, который получил широкое распространение в странах бывшего СССР, и амантадина, более известного в США.
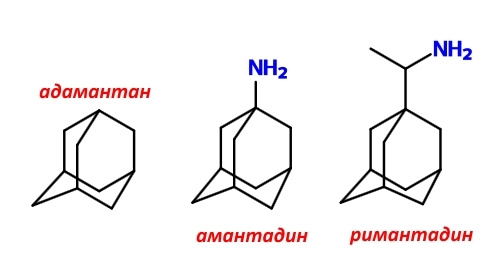
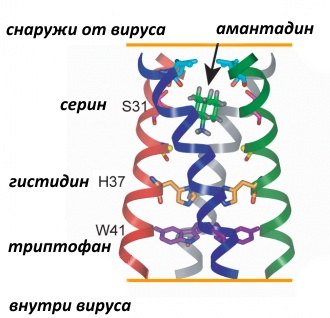
Эти соединения одновременно содержат фрагменты каркасного углеводорода адамантана, которые могут выступать заглушкой канала, образованного M2-белком, а также аминогруппы, которые позволяют удерживаться водородными и/или электростатическими связями (после протонирования) с соответствующими аминокислотными остатками.
Отличительной чертой многих РНК-вирусов, к которым принадлежит и вирус гриппа А, является высокая частота мутаций. Если вы помните, так называемый свиной грипп H1N1, который пришёл из Мексики в 2009 и уже давно стал частью циркулирующих сезонных вирусов, устойчив к производным адамантана из-за мутаций в трансмембранном сегменте белка M2. Так что более назначение этих лекарств при гриппе не рекомендуется.
С помощью ядерного магнитного резонанса установлено, что производные адамантана внутри канала застревают на уровне остатков аминокислоты серина (S31 на рисунке выше). Однако очень часто в результате мутации остаток серина заменяется на аспарагин. При этом, как полагают, диаметр канала увеличивается, и молекула лекарственного средства уже не может прочно в нём удерживаться, таким образом канал остаётся заблокированным меньшее время, что позволяет вирусу делать своё чёрное дело.
В настоящее время ведутся разработки соединений, который бы могли затыкать или иным образом нарушать работу М2-канала у мутировавших штаммов вирусов, однако, несмотря на обилие публикаций, о клинических испытаниях подобного рода соединений мне не известно.
Продолжение следует.
Вопросы для самоконтроля. Ну первое сентября же!
1. Почему растительные вирусы не имеют суперкапсида?
2. Какие преимущества даёт вирусу возможность связывания с несколькими рецепторами?
3. Известно, что ВИЧ-1 может связываться с 2 корецепторами: CCR5 и СXCR4. Как этот факт может сказаться на эффективности препарата маравирок?
4. Каким образом может сказаться на эффективности препарата маравирок замена в его молекуле 2 атомов фтора на 2 атома хлора? Поясните свой ответ.
5. Энфувертид вводится в организм человека только инъекционно. Как, по-вашему, с чем это связано?
6. В литературе встречается следующее неправильное объяснение механизма действия римантадина и амантадина: указывается, что эти препараты блокируют
закисление эндосом, поскольку являются веществами основной природы (аминами). Какой описанный в статье экспериментальный факт опровергает такое объяснение?
7. Какую модификацию молекул ремантадина и амантадина вы бы предложили провести, чтобы их производные смогли действовать на S31N мутанты?
Источник


