Термин «живое ископаемое» был впервые введен Чарльзом Дарвином в его фундаментальном труде «О происхождении видов» в 1859 году. Ученый тогда обратил внимание на анатомическое строение утконосов и лепидосирен (вид двоякодышащих рыб). Он предположил, что эти животные в процессе эволюции морфологически изменяются очень медленно, ввиду отсутствия сильной конкуренции и резких изменений в окружающей среде.

Тем самым ученый пришел к выводу, что новые формы не успевают быстро образовываться, а старые формы быстро вымирать и, поскольку анатомически лепидосирены и утконосы напоминают давно вымерших животных из своих отрядов, он рискнул назвать этих представителей «живых аномалий» «почти ископаемыми» .
С тех пор этот термин стал иметь двоякий смысл. С одной стороны, живыми ископаемыми стали называть виды, которые не имеют живых близких родственников уже давно, с другой стороны — виды, практически не изменившиеся в процессе эволюции. Однако Дарвин использовал этот термин только в двух проходных комментариях, а популяризировали его уже спустя восемьдесят лет после открытия коморской латимерии.
Коморская латимерия (Latimeria chalumnae) — вид рыбы из рода латимерия, открытый в 1938 году и отнесенный к отряду целакантообразных. В народе их ошибочно называют «целакантами». Последние же на самом деле относятся к другому роду и к другому семейству, которые в свою очередь являются вымершими.

Ошибочное представление о латимериях связано с тем, что представителей данного рода долгое время считали редкими и вымирающими видами «древних существ», которые якобы практически не изменились за последние 400 миллионов лет своего существования.

Такой подход в конце концов привёл к переносу концепции «живой» окаменелости на «ДНК», в результате чего появилась сомнительная гипотеза о так называемом генетическом застое, который в корне противоречит принципам эволюционной генетики. Согласно этим принципам геномы в среде постоянно изменяются под совокупностью воздействий различных мутационных процессов, образующих новые варианты генетических последовательностей, которые вследствие генетического дрейфа и отбора либо устраняются, либо закрепляются в популяциях. Т.е. даже при стабилизирующем отборе, накопление нейтральных мутаций в поколении так или иначе меняют геном, ведь суть отбора заключается не в сохранении особей совершенно неизменными в стабильных условиях, а сохранение особей стабильно меняющихся в пользу эволюции среднего признака, против особей имеющих крайние отклонения от средней нормы.
Это в свою очередь означает то, что единственная возможность для геномов реплицироваться без изменений будет подразумевать как минимум одно из двух следующих условий:
-
новые варианты не появляются (т.е. нет никаких мутаций)
-
новые варианты систематически устраняются путем отбора (т.е. без генетического переноса и очень сильного отбора против новых вариантов.)
Однако, несмотря на абсурдность данной гипотезы, ряд учёных провели исследование, в котором отметили бесспорность замедленной генетической эволюции, основываясь всего на трёх ядерных генах и двух целых митохондриальных геномах, однако отметив при этом и дивергенцию популяций латимерий, которая при замедленной эволюции почему-то активно происходит и по сей день.
Мы должны понимать, что геном это не несколько ядерных генов и не две последовательности мт. генов цитохром-с-оксидазы. Конечно существуют консервативные гены, которые практически не меняются и так называемые консервативные митохондриальные геномы, чья степень консервативности измеряется в количестве и скорости замен в них происходящих, по отношению к другим группам живых организмов. Однако нужно учитывать, что а) — скорость мутаций может быть неравномерной и может вполне различаться для разных видов живых организмов; б) некоторое количество консервативных генов не сделают Вас живым ископаемым, если все остальные гены «мутировали» постоянно.
Устранить недоразумение о генетическом застое и опровергнуть подобные «бесспорные» заявления вызвался профессор лаборатории Эволюции, Геномов, Поведения и Экологии парижского университета Национального Центра Научных исследований Дидье Казан. В своём исследовании он заново оценил темпы геномной и анатомической эволюции, рассмотрев имеющиеся молекулярные и морфологические данные для этой группы животных. Наиболее ярким его примером, доказывающим отсутствие замедленной эволюции, является филогенетическое исследование сорока четырёх ядерных генов, которое не показывает резкого снижения скорости молекулярной эволюции в линии «целакантов». Более того: биология «целакантов», по мнению учёного, не предполагает никаких очевидных причин, по которым они вдруг «решат» эволюционировать медленно.
Они не живут в среде, которая предполагает направленный отбор на репликацию и репарацию ДНК, что приводит к очень низкой частоте мутаций. У них нет чрезвычайно долгого времени генерации и они определённо не образуют больших популяций. Кроме того, хорошо известно, что скорость замещения генов изменяется в геноме в зависимости от нескольких факторов, таких, как локальная скорость рекомбинации и расстояние до источника репликации. Также известно, что скорость замещения непостоянна в разных линиях. Ко всему прочему учёный также отметил роль аминокислотных взаимодействий, которая, вероятно, является важной в изменении скорости эволюции белка в различных линиях. Именно поэтому при сравнении генов разным исследователям может показаться, что некоторые гены эволюционируют быстрее, а другие наоборот – медленнее. Исходя из этого он подчеркнул, что, даже если бы действительно существовал генетический застой в масштабе всего генома, маловероятно, что он мог бы объяснить простым способом скорость морфологической эволюции, поскольку медленная генетическая эволюция не связана со скоростью последней в разных линиях этих рыб, к тому же морфологического стазиса у латимерии попросту не обнаруживается.
В геологической летописи нет окаменелостей, идентичных двум живым видам латимерии. И нет окаменелостей, которые можно было бы отнести к роду Latimeria. Это наводит на мысль, что палеонтологи – даже те, кто убеждён, что современные целакантообразные являются «живыми ископаемыми» – считают морфологические различия между дожившими до нас «целакантами» и ископаемыми настоящими целакантами настолько обширными, что их следует сгруппировать в отдельные роды.
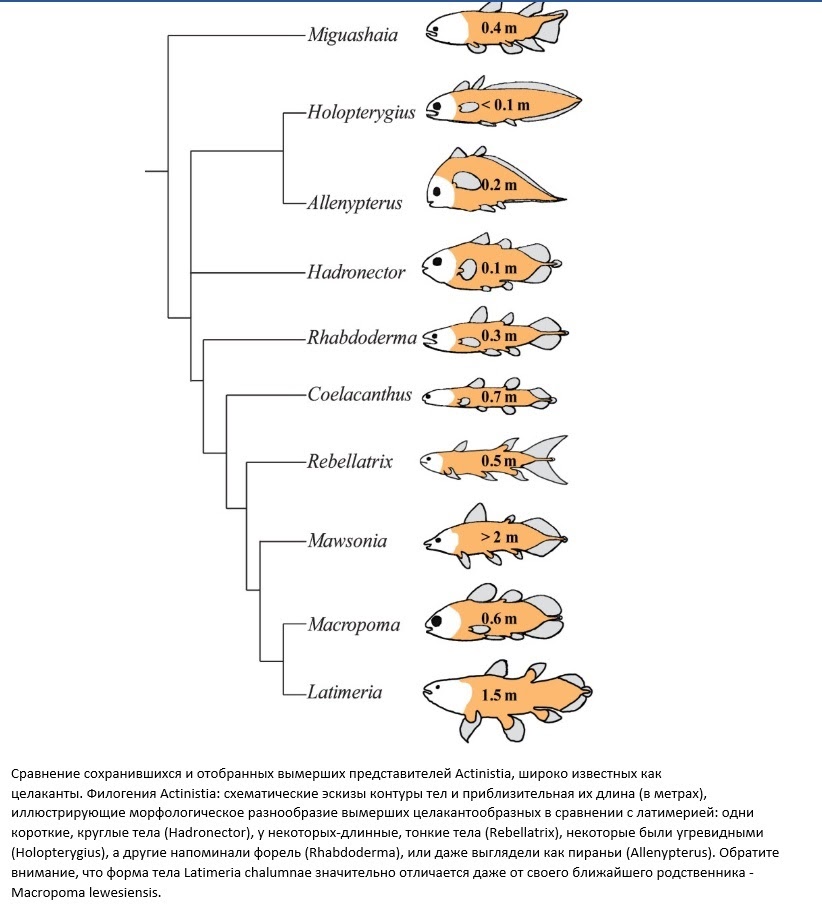
Помимо этого, против «живых ископаемых», говорит и тот факт, что даже самый близкий вымерший родственник латимерии из сестринского рода Macropoma имеет ряд некоторых интересных отличий. Мало того, что вымерший родственник в три раза меньше своего двоюродного брата (полметра против полутора метров), так он ещё и имеет другую внутреннюю морфологию. Так, например, плавательный пузырь в макропоме окостенел (на рисунке ниже посмотрите на «В» — пузырь ярко бросается в глаза), а у латимерии он напротив заполнен жиром, что указывает на то, что макропома, вероятно, жила в другой среде. Также на это указывают и существенные различия в позвоночном столбе (постанальная область короче, а вентральные шипы расширяются менее вентрально). У макропомы по-другому устроены плавники и совсем иначе устроена анатомия черепа, из-за чего, собственно, у неё весьма сильно отличается морфология головы.
 с плавниками и черепом её ближайшего родственника (справа)")
Все эти факты предельно намекают на то, что при всей внешней морфологической схожести, основанной на четырёх мясистых лопастных плавниках, разные целакантообразные, вероятнее всего, будут иметь разную внутреннюю организацию тела. Даже у современных целакантообразных в разных популяциях рыб будут по-разному выглядеть позвоночные столбы, демонстрируя значительные вариации в количестве нервных клеток. В общем и целом всем тем, кто считал, что все «целаканты» «на одно лицо», латимерия даёт пощёчину своим «целакантовским» хвостом!
В течение последнего столетия понятие «живое ископаемое» как результат эволюционного застоя всё чаще рассматривается как вводящее в заблуждение, поскольку оно само по себе, как и его синонимы по типу — «базальное происхождение» или «примитивные сохранившиеся виды», не имеют смысла даже в построении классификации, не говоря уже о самой точке зрения древовидного мышления. Клада с людьми и клада с латимериями расходились в одном узле и, стало быть, они имеют одного последнего общего предка. И если кладу с Homo «пропустить» через выборку традиционной филогении, поставив Homo в качестве единственного члена клады Rhipidistia и включив в выборку всех ископаемых актинистов, то мы увидим, что род Latimeria, как и любые другие современные роды принадлежат к большой кустистой кладе, где все ископаемые окажутся где-то далеко в начале, а не ископаемые в конце. Когда Дарвин вводил термин «живое ископаемое», маловероятно, что он действительно думал, что существующий вид будет идентичен предковому виду. Вероятно, это уже додумали за самого Дарвина. Как следствие, идея о том, что латимерия является «живым ископаемым» есть ложная вера, искажающая правду, хотя для воспаленного разума криптозоолога, креациониста и конспиролога всё будет ровным счётом наоборот , ведь учёные всё врут и скрывают. Однако это уже совсем другая история.
Примечание
Материал был написан мной и опубликован в научно-популярном сообществе фанерозой.
Источник:
Insights & Perspective :Why coelacanths are not ‘living fossils’
A review of molecular and morphological data
Didier Casane and Patrick Laurenti; 2013


