
В работе над симулятором нервной системы пока мы касались только с хорошо изученными аспектами её работы. Но сложность моделирования нервной системы и причина, по которой до сих пор не создан искусственный интеллект – это отсутствие полного представления о том, как работает нервная клетка. Подробно описаны многие процессы, протекающие в нервной клетке и нервной системы в целом, но нет четкого алгоритма их работы, который можно было бы перенести в модель или компьютерную программу.
Простая идея алгоритма работы нейрона позволила решить эту проблему.
2. Симулятор нервной системы. Часть 2. Модулируемый нейроэлемент
3. Симулятор нервной системы. Часть 3. Ассоциативный нейроэлемент
4. Память, консолидация памяти и бабушкины нейроны
5. Моделирование эмоций или электронное чувство новизны
6. Удивительный мозжечок
7. Структура и стартовые настройки мозга
Мне нравится аналогия о составлении теории работы мозга с собиранием пазла из книги Джеффа Хокинса «Об интеллекте». При составлении этого пазла у нас отсутствуют некоторые элементы, а некоторые элементы от другой головоломки, но мы имеем большой массив данных о нервной системе и мозге, что значит у нас в наличии почти собранная головоломка, поэтому мы можем примерно представить всю картину, и, используя наше воображение определить недостающие элементы.
Моя цель создание логической модели работы нервной системы, можно сказать создание наброска того, что изображено на незаконченном пазле, причем он должен соответствовать и не противоречить всем имеющимся элементам головоломки и при этом быть логически законченным. Чтобы заполнить пробелы была создана некоторая теоретическая основа, которая возможно некоторым покажется спорной. Но для модели на данном этапе, главное это то, что она позволяет эмулировать как внутренние, так и внешние наблюдаемые явления, протекающие в нервной системе. В рамках полученной модели возможно объяснение многих явлений, таких как память и консолидация памяти, эмоции, специализация нейронов и многое другое.
Во второй части мы выяснили, что существуют три типа рефлекторной деятельности установленной академиком И.П. Павловым. Если с биологическими механизмами привыкания и сенсибилизации всё предельно ясно, то с образование условных рефлексов не все так просто как кажется. Дело в том, что широко изучены и описаны внешние проявления этого механизма, нет объяснения того как это происходит на клеточном уровне.


К примеру, мы знаем, что при сочетании активности двух нервных центров со временем между ними формируется рефлекторная дуга. Т.е. впоследствии при активации одного нервного центра будет, происходит передача возбуждения к другому нервному центру. Если мы образно разделим такую рефлекторную дугу на сегменты, и рассмотрим такие сегменты как отдельные элементы. То можно говорить, что при образовании рефлекторной дуги условного рефлекса в каждом сегменте происходит коммутация направленного характера. Каждый сегмент выбирает определенное направление, в котором будет, происходит передача нервного возбуждения при его активации. Конечно, стоит отметить, что данное направление не определено четко для сегмента, а может коррелироваться в определенных значениях. Даже можно говорить об усилении передачи в определенном направлении и ослаблении в других направлениях.

При укреплении рефлекса многократными повторениями можно говорить об уточнении и усилении передачи в направлении для каждого сегмента. Эта концепция приводит к выводам, что ели мы разделим всю кору на подобные сегменты, то будем наблюдать в каждом некую ориентацию по направлению с различной точностью и силой. Каждый сегмент будет, вялятся частью какой-то рефлекторной дуги условного или безусловного рефлекса. Предположительно эта ориентация в процессе обучения может уточняться или меняться.

Если обратится к нейронной парадигме, то в ней не предусматривается ориентация по направлению. У нас есть мембрана и дендриты, принимающие сигналы и аксон, по которому сигнал передается далее к другим клеткам после пространственно-временной суммации, то есть сигнал передается в одном направлении по аксону к его окончаниям. Но при этом мы все равно наблюдаем формирование направленного распространения возбуждения в мозге, при образовании условных рефлексов.
Если рассматривать не отдельную клетку как функциональную единицу направленной коммутации, а небольшую область клеток, то можно увидеть, что клетки и их отростки очень плотно переплетены, причём в разных направлениях. Это дает элемент направленной коммуникации с множеством входов и выходов в различных направлениях.

Форма нейрона обусловлена эволюционными изменениями. Форма клетки сформировалась в нервных системах, в которых осуществлялся только простейший функционал нервной деятельности. Когда же развитие жизни на Земле потребовало добавить в набор функций нервной системы образование уловных рефлексов, то эволюция пошла по пути не перестройки клетки, а увеличению их количества и плотному переплетению их отростков.
Таким образом, свойство направленной коммутации распределено в группах нейронов, в изменении силы их синапсов. Ассоциативный нейроэлемент является в моделировании функциональной единицей и поэтому аналогом в биологии для него является группа нейронов, для которой будет выражено явление направленной коммутации.
Мы выяснили, что для нас важно направление распространение возбуждения, но как происходит определение этого направления для каждого функционального элемента. Известно, что возбуждение стремится распространиться к другому источнику возбуждения, причем более сильный и масштабный очаг возбуждения притягивает к себе более слабые (заключение Павлова И.П.). Т.е. если функциональный элемент получит возбуждение, то каким-то образом он должен определить направление, которое впоследствии сформируется и сохранится в его структуре.
В своей работе по моделированию я отталкивался от идеи электромагнитного взаимодействия нервных клеток, и эта идея дала ответы на множество загадок о мозге, дала теорию и модель, объясняющую многие аспекты работы нервной системы.
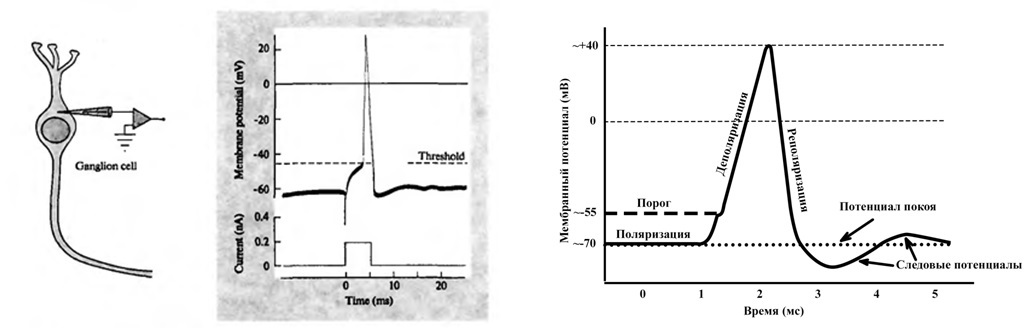
Нервный импульс во всей нервной системе имеет одинаковую форму, и по аналогии с ним ассоциативный нейроэлемент имеет свойство заряд характеризующее изменение совокупного заряда на поверхности мембран функциональной единицы. Т.е. задан определенный закон изменения некоторой характеристики именуемой заряд.
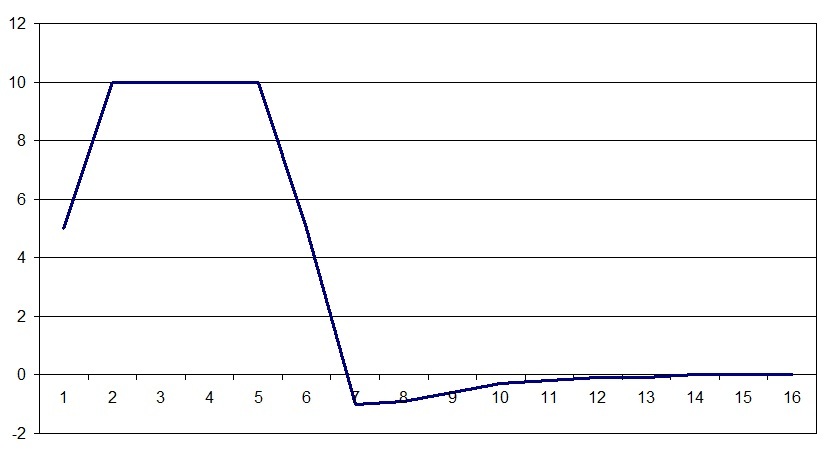
Так задан закон в программе, шкала по горизонтали это время в сотых секунды, по вертикали заряд в относительных единицах. Он несколько отличается от графика спайка тем, что часть максимума более продолжительна по времени. Это связано с тем, что значения спайка определяются в одной точке нервной ткани при прохождении возбуждения, а график заряда это отражение заряда по всех поверхности клетки или группы клеток. Так же за ноль по шкале заряда принято состояние покоя нервной ткани. Следует отметить, что закон изменения заряда так же отражает следовой потенциал, который ранее считался следствием некого колебания или выравнивания зарядов разделенных мембраной, но для модели такое поведение заряда оказалось очень важным.

На рисунке выше представлена схема ассоциативного нейроэлемента. Сигналы от синапсов прямого действия (X1, X2, X3 … Xn) поступают в сумматор (а). И если результирующая сумма превысит определенный порог (б), то произойдет активация нейроэлемента. При активации нейроэлемента его заряд начнет изменяться в соответствии с установленным законом (в). Информация об этих изменениях и местоположения самого элемента будет доступна всей системе. Далее в определенный момент времени запускается механизм определения вектора предпочитаемого направления распространения возбуждения (г). Это происходит путем получения некого среднего положения заряда всех активных нейроэлементов, т.е. центра масс зарядов, характеризуемой точкой в пространстве. Данную точку назовем точкой паттерна, потому что для каждой комбинации активных клеток и состояния их зарядов в вычисляемый момент времени для каждого нейроэлемента, положение этой точки будет своё. Проще говоря, заряды нейроэлементов влияют на определение вектора направления предпочитаемого распространения возбуждения, положительный заряд притягивает возбуждение, отрицательный заряд отталкивает.
Для определения вектора предпочитаемого распространения возбуждения подобрано правило:

где r вектор начало которого находится в центре нейроэлемента для которого определяется вектор, а конец в центре n-ного нейроэлемента.
Правило и закон изменения заряда подбирался эмпирическим путем, так чтобы имитировать образование условных рефлексов. Подробнее в статье.

После получения вектора предпочитаемого направления распространения возбуждения (T), происходит вычисления силы синапсов (Y1, Y2, Y3 … Yn). Каждый синапс охарактеризован вектором синапса (S), начало которого лежит в центре нейроэлемента а конец связан с центром целевого нейроэлемента на который осуществляется передача сигнала. Основной параметр синапса это его сила F, значение силы ограниченно в определенных рамках, к примеру, побудительный синапс может иметь значения от 0 до 10.
Представим, что вектор Т формирует вокруг себя некий конус вершина которого находится в центре нейроэлемента, а плоскость основания перпендикулярна вектору T, если вектор синапса попадает в область ограниченную данным конусом, то значение силы синапса будет увеличено на определенное значение. И соответственно, если вектор синапса оказывается за пределами области конуса, то происходит уменьшение силы синапса, но при этом значения силы не выходит за пределы установленного максимума и минимума.
Область конуса вокруг вектора T охарактеризована углом при вершине данного конуса, этот угол называется фокус. Чем меньше фокус, тем точнее будет определено направление передачи возбуждения в нейроэлементе. Как говорилось ранее при повторении организмом одного и того же условного рефлекса происходит его уточнение. Поэтому для модели был выбран следующий метод изменения фокуса, при вычислении вектора Т происходит сравнение его с предыдущим его значением, и если вектор измен незначительно то фокус уменьшается на некоторое значение, но если вектор был изменён сильно, то фокус возвращается к своему максимальному значению. Это приводит к постепенному уменьшению фокуса при многократном повторении одних и тех же условий.
Здесь очень важный момент, это то, насколько будет изменяться сила синапсов при каждой активации. Это определяется параметром нейропластичность P.
Формула нового значение силы синапса, будет иметь вид:
Fnew = Fold + I × P × (Fmax — Fmin);
Fmin ≥ Fnew ≥ Fmax;
где P — нейропластичность (0 ≥ P ≥ 1);
I – параметр определяющий находится ли вектор синапса в пределах области повышения силы синапса (I = 1) или в области понижения силы синапса (I = -1);
Fold – предыдущее значение силы синапса;
Fmin – минимальное значение силы синапса;
Fmax – максимальное значение силы синапса.
Нейропластичность в биологии характеризует то, насколько нейрон податлив к изменениям своей структуры под влиянием внешних условий. Для различных областей мозга характерна своя степень пластичности, так же она может изменяться в зависимости от некоторых факторов.

Это пример позволяет понять, как на основе ассоциативных нейроэлементов формируются условные рефлексы. Белые нейроэлементы формируют рефлекторную дугу безусловного рефлекса с заголовком “R” и ответом “1”. Эти нейроэлементы не изменяют значений сил своих синапсов. Синие нейроэлементы не участвуют изначально ни в каких рефлекторных актах, они как бы заполняют все остальное пространство нервной системы, и они случайно связаны между собой посредством синапсов. Поэтому если мы будем активировать один такой нейроэлемент связанный с рецептором “Q”, то возникнет некоторый очаг возбуждения имеющий случайный характер распространения и зациклившись сам на себя через некоторое время он потухнет, не создав никого ответа. Если мы будем сочетать примерно в одинаковом временном интервале безусловный рефлекс с заголовком “R” и активацию рецептора “Q”, то сформируется рефлекторная дуга условного рефлекса. И уже активация просто рецептора “Q” будет приводить к ответу “1”.
Для наглядности и оптимизации работы модели было применено динамическое создание нейроэлементов, которое эмулирует заполненное пространство нервной системы случайно связанными между собой элементами. Здесь не моделируется какой-то рост новых нейронов, или новых связей, все изменения происходят только в силе синапсов, просто нейроэлементы ранее не вовлеченные в какой либо рефлекторный акт не показаны.

Следующий пример показывает, как ведет себя возбуждения при активации различных центров при равных условиях и при абсолютной пластичности (P = 1).
Изменение направления распространения возбуждения под влиянием двух центров возбуждения, когда пластичность абсолютна (P = 1):
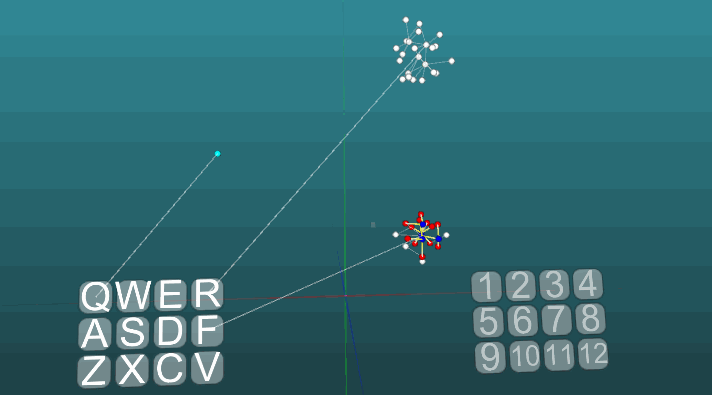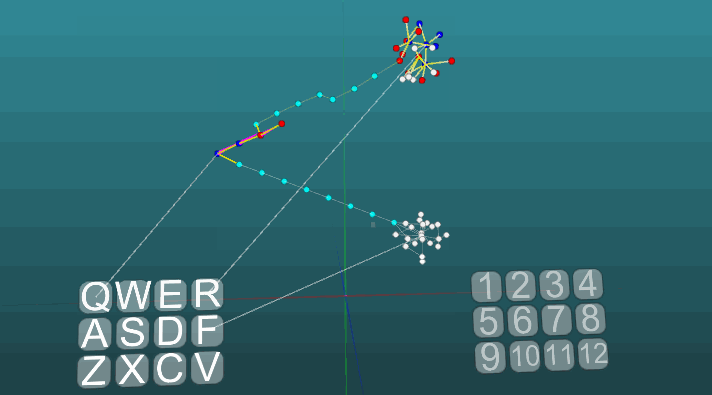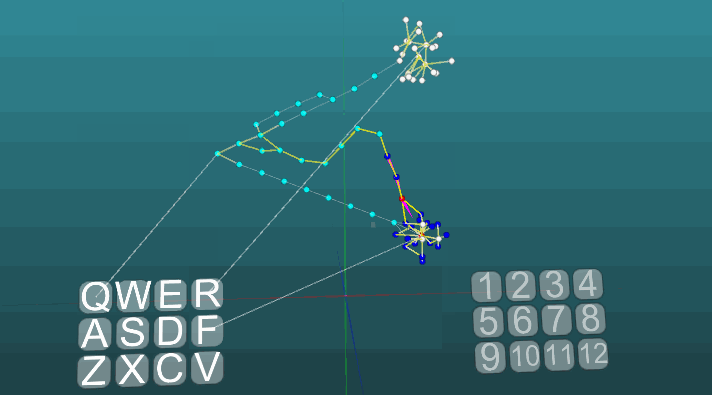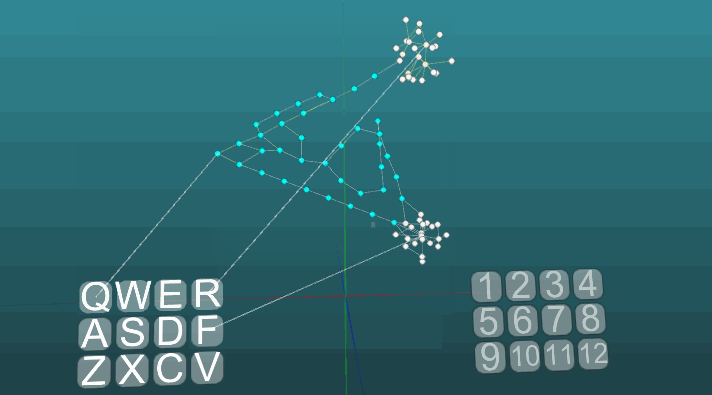
И при низкой пластичности (P = 0.1):
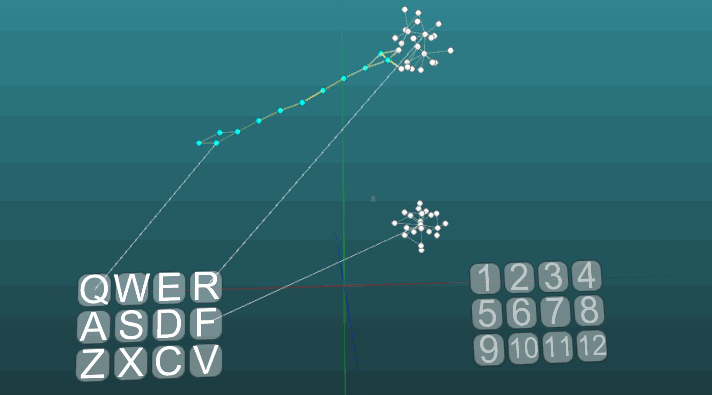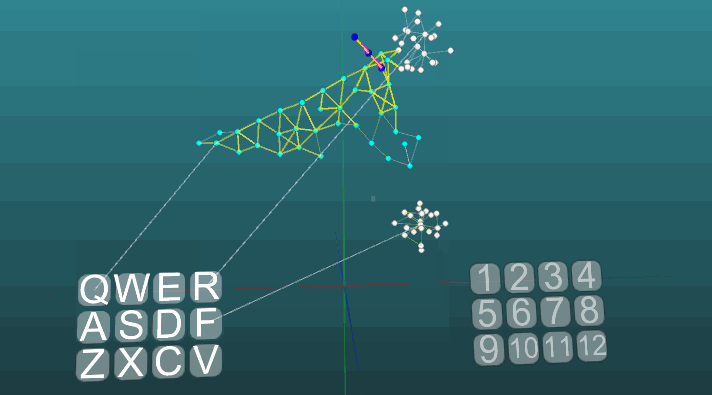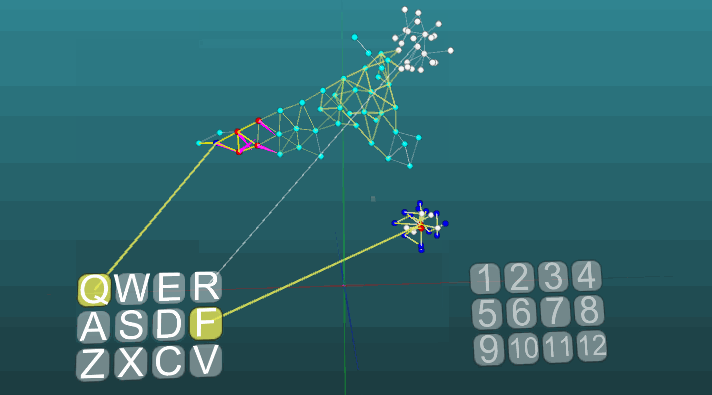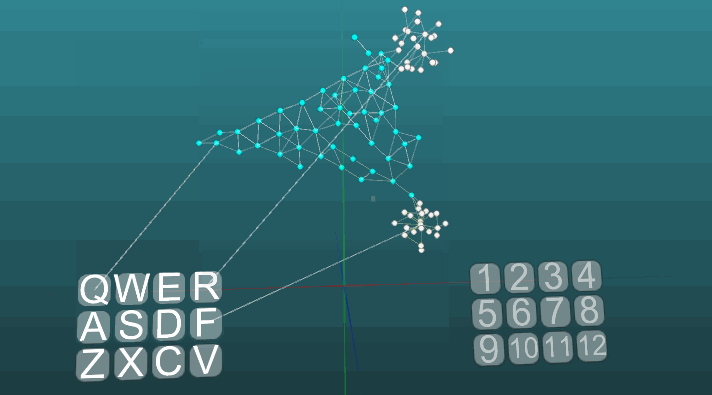
На этом мы закончили рассматривать основы модели нервной системы. В следующей части мы рассмотрим прикладные вещи, как все это использовать, что бы имитировать память, эмоции, специализацию нейронов.

Источник

