
К паукам можно относиться как угодно: их можно бояться, недолюбливать или держать в качестве питомцев. Но любой, от арахнофоба до арахнолога, согласиться с тем, что они мастера по строительству своих сетей. Научное сообщество уже очень давно и с большим интересом наблюдает за членистоногими прядильщиками, но полностью раскрыть все их секреты пока еще не удалось. И вот ученые из университета Джонса Хопкинса (США) решили детально рассмотреть и описать процесс строительства паутины, используя при этом искусственный интеллект и приборы ночного видения. Выяснилось, что разные виды пауков подчиняются общим правилам в ходе создания своих сетей. Следовательно, наблюдение за движениями лапок может предсказать, что именно будет строить паук. На какие стадии можно разделить строительство паутины, как пауки ведут себя во время каждой из них, и как эти данные могут помочь в понимании нас самих? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Пауки — это удивительные создания, какими бы страшными они не казались для некоторых людей. Терпеливые и коварные охотники, талантливые строители — все эти по большей степени нелестные эпитеты тем или иным образом отображает поведение пауков: метод добычи пищи, наличие паутины и/или яда и т.д.

Картина «Минерва и Арахна» (Рене-Антуан Уасс, 1706 год).
Если вспоминать древнегреческую мифологию, то самым ярким появлением пауков в культурной среде человека является история об Арахне. Она была невероятно талантливой ткачихой, решившая, что сможет превзойти Афину. И ее уверенность в себе была не безосновательна, ибо ей удалось победить в ткацком состязании богиню. Однако Афина, которая хоть и была богиней мудрости, но уж точно не терпения или умения проигрывать, впала в ярость, разорвала работу Арахны и ударила ее. Для Арахны это было невероятным унижением, и она повесилась. История не очень позитивная, но дальше больше. Афина, решив, что смерть это недостаточное наказание, воскресила Арахну, превратила ее в паука и обрекла вечно прясть.
Для нас этот мир говорит о двух вещах. Во-первых, древнегреческие боги далеки от идеала. Во-вторых, пауки присутствовали в культуре человека многие тысячи лет и занимали там далеко не самое последнее место. Подобное про пауков можно сказать и в аспекте других культур и религий.

Реальный облик цутигумо.
Цутигумо (土蜘蛛) — паукоподобные ёкай (сверхъестественные существа) в японском фольклоре. В одной из легенд героя по имени Минамото-но Райко заманивает в дом и помещает в иллюзию цутигумо в облике мальчика. Но герой разрушает иллюзию, ударив малька мечом. Очнувшись от дурмана, он обнаруживает, что опутан паутиной, а мальчик на самом деле это огромный паук.

Реальный облик Греты.
Вполне вероятно, что персонаж Грета из сериала «Love, Death & Robots» (серия №7 «Beyond the Aquila Rift») основан именно на цутигумо (как по мне, аналогия на лицо).
От культуры перейдем к науке. Факт того, что паучий шелк обладает невероятной прочностью и эластичностью, известен уже давно. Существует масса исследований, в которых описываются методы искусственного воссоздания паучьего шелка с последующим его применением в самых разных областях, от медицины до космонавтики.
Но в рассматриваемом нами сегодня труде ученые уделили внимание не столько самой паутине и ее составу, сколько тому, как именно пауки создают из нее свои сети.
Ученые отмечают, что внутренние состояния, такие как сытость, сексуальное возбуждение или агрессия, сильно влияют на поведение любого животного. Однако проблема для многих поведенческих парадигм состоит в том, что эти состояния должны выводиться косвенно, часто на основе стохастического переключения состояний поведения. Другими словами, крайне часто у этих поведенческих аспектов и переходов между ними нет физической репрезентации, которую можно было бы оценить. К примеру, чувство голода. Обычно, голодное животное больше двигается, т.е ищет пропитания, а сытое — остается на месте (отдыхает). Но сытые животные тоже могут бродить по лесу (джунглям, саваннам и т.д.), а голодные лежать и бездельничать. Другими словами, такой метод (т.е. эмпирический) поведенческого распределения крайне непоказателен. Чего не скажешь о пауках.
Преимущество количественной оценки поведения пауков-кругопрядов (которые делают спиральные сети в форме колеса) состоит в том, что построение сети разделяется на поведенческие фазы, которые легко определяются траекторией паука и геометрией паутины.
Любопытно и то, что многие виды пауков из семейства Araneidae и Uloboridae разделяют одну и ту же последовательность этапов построения паутины, которые являются результатом многих общих форм поведения, таких как ходьба и экструзия шелка. Хоть некоторые из этих поведений и проявляются на всех этапах создания сети, то, как эти поведения координируются, различается на разных его этапах.
В принципе, различные фазы построения сети должны быть предсказуемы на основе этих различных поведенческих стратегий, независимо от траектории центроида паука. Несмотря на то, что последовательность этапов создания паутины довольно однотипна, часто наблюдались так называемые атипичные паутины. Такие атипичные сети и процесс их создания называют «сенильными» (хлипкими, если проще). В их случае изменяются особенности и регулярность геометрии сети, а также бывают случаи прерывистых и повторяющихся этапов построения, которые необязательно приводят к изменению конечного состояния паутины. Однако, изменения в поведении, которые приводят к таким атипичным сетям, фактически не изучены.
В данном труде ученые демонтируют аналитическую основу для определения поведения в ходе строительства сетей, за счет чего удалось выявить однотипные (характерные) последовательности действий, которые характеризуют построение паутины.
Результаты исследования
Большинство пауков-кругопрядов в основном активны весной и летом, но другие виды, такие как Uloborus diversus, активны в течение всего года. Из-за этой продолжительной активности, а также из-за других особенностей, таких как их небольшой размер и толерантность к сородичам, было решено использовать для исследований именно их (1А).
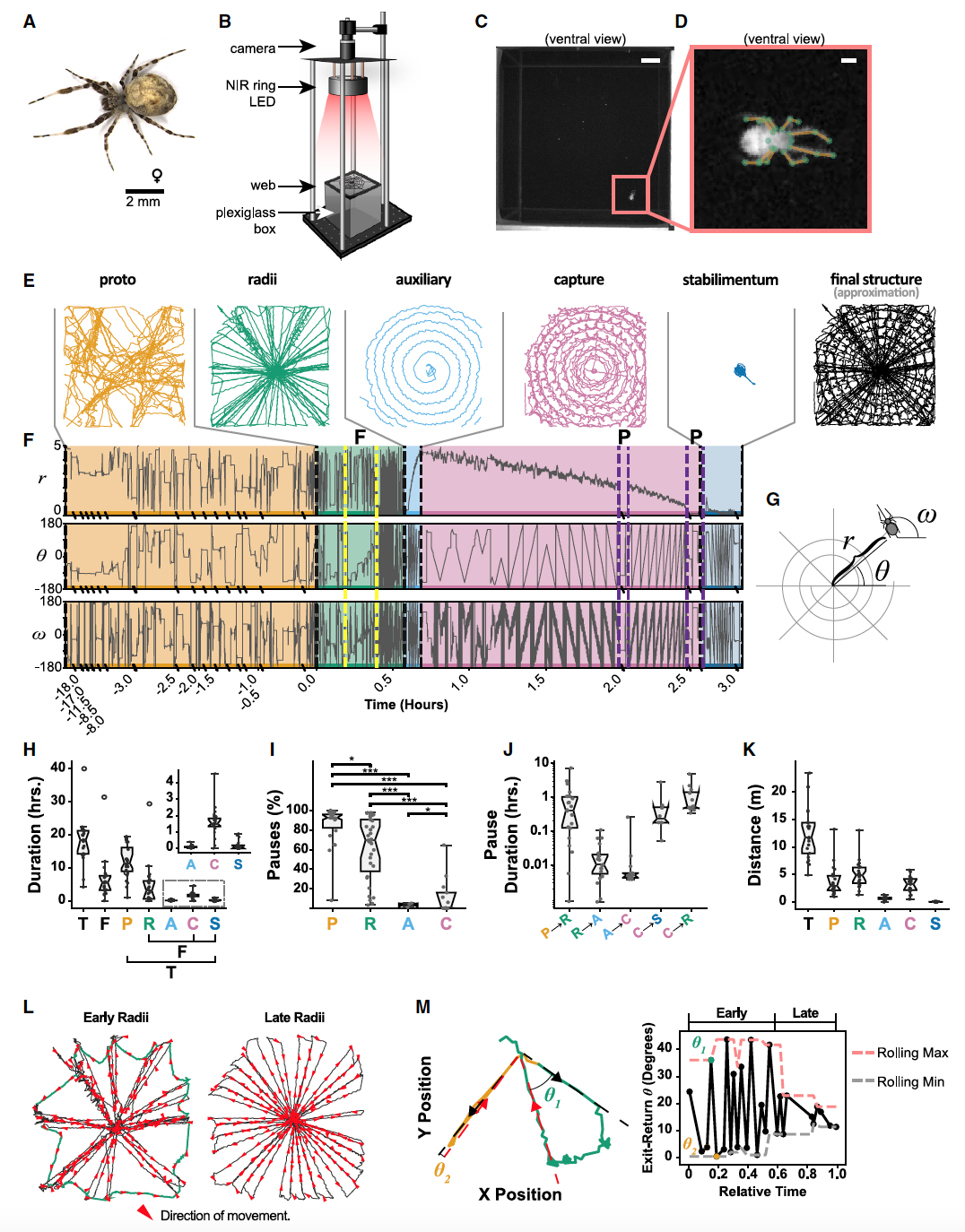
Изображение №1
Данный вид может строить паутину внутри самой разной геометрии, но предпочитает создавать горизонтальные сети, делая это в ночное время суток. Эти особенности были учтены при создании камеры для наблюдений (1B—1D).
Поведение паука регистрировалось с частотой кадров 50 Гц в течение 24 часов, тем самым охватывая все этапы строительства сети. Одно лишь визуальное наблюдение показало наличие общей траектории, которую используют большинство пауков-кругопрядов (1E—1H).
Как и в случае с большинством пауков-кругопрядов, типичное развитие плетения сети у U. diversus начинается с прото-паутина, за которой следует построение радиусов и каркаса (внешней оправы), затем вспомогательная спираль, за которой следует спираль захвата. У U. diversus, как и у некоторых других его сородичей, имеется дополнительная особенность их паутины — «stabilimentum» (украшение, декорация), необходимая для маскировки паука или привлечения внимания добычи (1E и 1F).
Видео №1: траектории движения в паутине (соответствует 1E и 1F).
U. diversus тратит значительное количество времени на изучение арены и сначала строит неорганизованную сеть, называемую «прото-паутиной» (оранжевые линии на 1E и 1F). Эти сети не имеют очевидной регулярности, в большинстве случаев не являются частью окончательной структуры и построены с длинными нерегулярными паузами продолжительностью до 8 часов (1H—1K).
Предположительно, этот этап строительства связан с исследованием окружающей среды, когда паук оценивает структурную целостность своего окружения и определяет точки привязки для окончательной паутины. Этот этап часто заканчивался длительной паузой, которая могла длиться от нескольких минут до нескольких часов (1J).
Затем паук удалял большую часть прото-паутины, оставляя часть, необходимую для создания радиусов и каркаса (1L). Эта ранняя фаза построения радиусов часто включала перемещение паука наружу по радиусной линии и возвращение по вновь закрепленной (зеленые линии на 1M). Это характерно для построения новых радиусов по следующей тактике: закрепление шелка на ступице (хаб, точка соединения нескольких линий), движение наружу по предыдущему радиусу, затем закрепление нового шелка на каркасе на угловом расстоянии от предыдущей линии и возврат к ступице вдоль этой новой линии. Строительство каркаса часто является результатом такой последовательности: закрепление шелка на периферии перед возвращением, затем движение наружу по предшествующему радиусу, закрепление шелка каркаса в конце линии, а затем возврат к ступице (оранжевые линии на 1M). Эта ранняя фаза построения радиусов может быть определена путем расчета угла выхода и возврата из ступицы (1M), а также постепенного изменения углового положения паука (желтый контур на 1F).
На ранних стадиях строительства радиусов наблюдалась тенденция к чередованию траекторий выхода-возврата вдоль одной и той же линии с траекториями выхода-возврата, которые охватывали большой угол (1L и 1M).
Как только радиусы и каркас были готовы, паук ненадолго остановился, а затем разворачивался по спирали наружу от ступицы, чтобы создать вспомогательную спираль (синие линии на 1E и 1F). Этот этап длился всего несколько минут с несколькими паузами (1H — 1J). Этот этап необходим для стабилизации паутины для последующего строительства спирали захвата. Вспомогательная спираль носит временный характер, потому паук снимал ее по мере того, как продвигался внутрь от периферии, чтобы создать спираль захвата (розовые линии на 1E и 1F). В некоторых случаях после завершения спирали захвата паук добавлял «stabilimentum» (синие линии на 1E и 1F), после чего возвращался в центр готовой паутины, где мог неподвижно сидеть в течение нескольких дней в ожидании добычи.
Положения центурионов позволяют идентифицировать переходы между большинством этапов, но переход между прото-паутиной и радиусами кажется более постепенным и менее четким. Это не похоже на ограничение слежения за центроидом, а скорее отражает внутренне недискретную границу между этими этапами. На начальном этапе создания прото-паутины почти весь уложенный шелк в конечном итоге удаляется. Однако по мере продвижения этого этапа радиусы, которые будут частью окончательной структуры, создаются или изменяются, пока паук все еще удаляет прото-паутину. На этой ранней стадии построения радиусов паук часто пересекает радиусы несколько раз, регулируя полотно. Только когда прото-паутина была полностью удалена и каркас собран, паук сосредотачивался исключительно на конструкции радиусов и переходил к очень регулярной сборке дополнительных радиусов (1L).
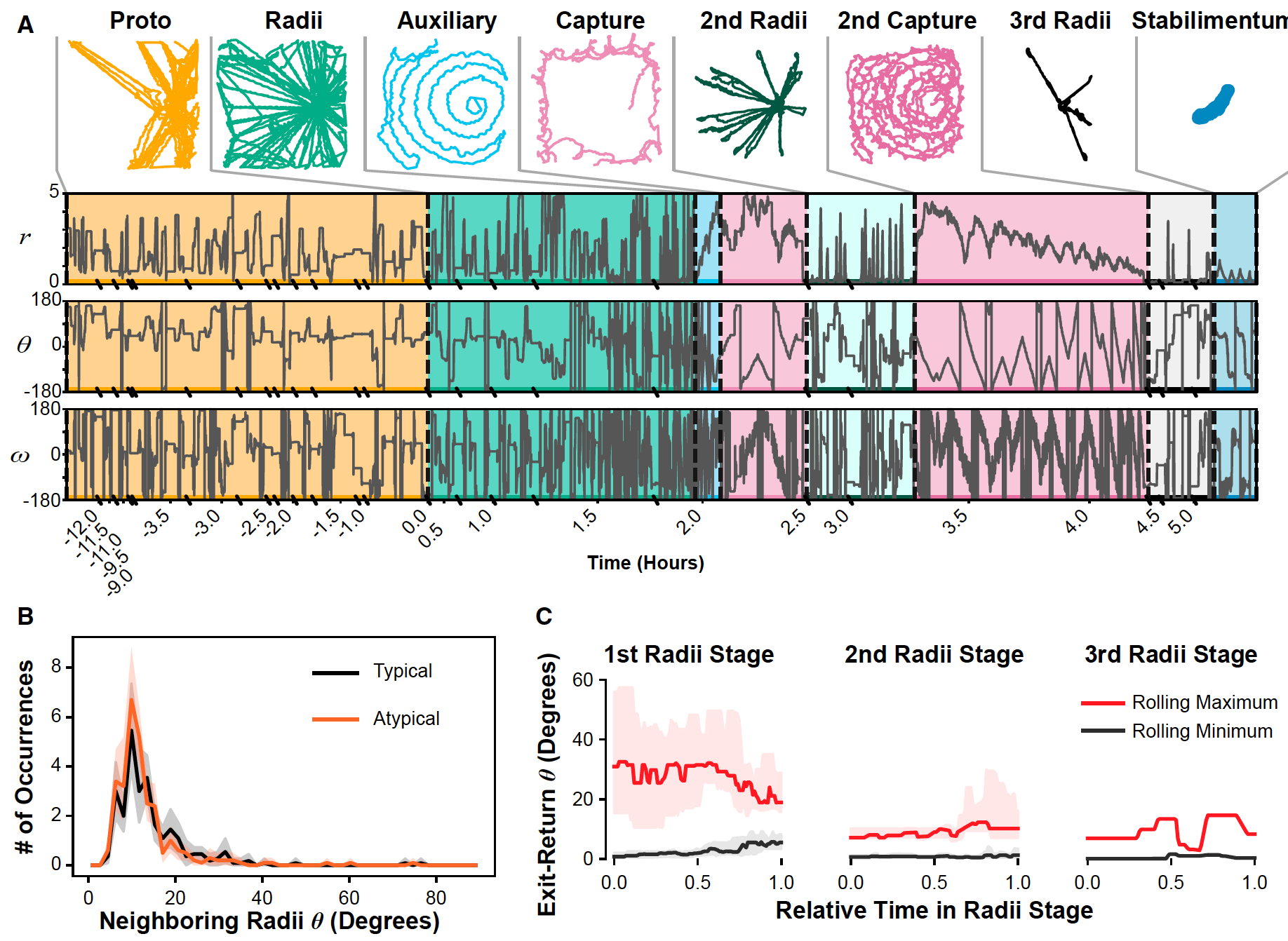
Изображение №2
Большинство пауков-кругопрядов следуют вышеописанной последовательности при создании своих сетей. Однако далеко не всегда все идет по однотипному сценарию — некоторые этапы меняются местами или вовсе пропускаются (2A).
Видео №2: траектории движения в паутине (соответствует 2A).
Любое отклонение от вышеописанной тактики строительства, по мнению ученых, следует именовать «атипичным». Из 21 сети, созданной пауками в ходе наблюдений, 11 можно было отнести именно к атипичным: этап создания спирали захвата дважды прерывался этапом радиального исследования, а в 9 случаях за созданием спирали захвата или размещением украшения следовал этап конструирования дополнительных радиусов.
Любопытно то, что дополнительный этап создания радиусов не был вызван нехваткой оных, так как плотность радиусов в начале этих атипичных стадий не отличалась от таковой в случае классической последовательности создания сети (2B). Другими словами, дополнительные радиусы были действительно дополнительными, а не попыткой перекрыть недостачу, возникшую на предыдущих этапах строительства. Но иногда этап дополнительных радиусов был связан с заменой старых на новые.
Большинство отклонений от этапа создания спирали захвата оказались исследовательскими, так как радиальные траектории пауков следовали предшествующим траекториям, что указывает на то, что они шли по ранее построенным линиям (2С).
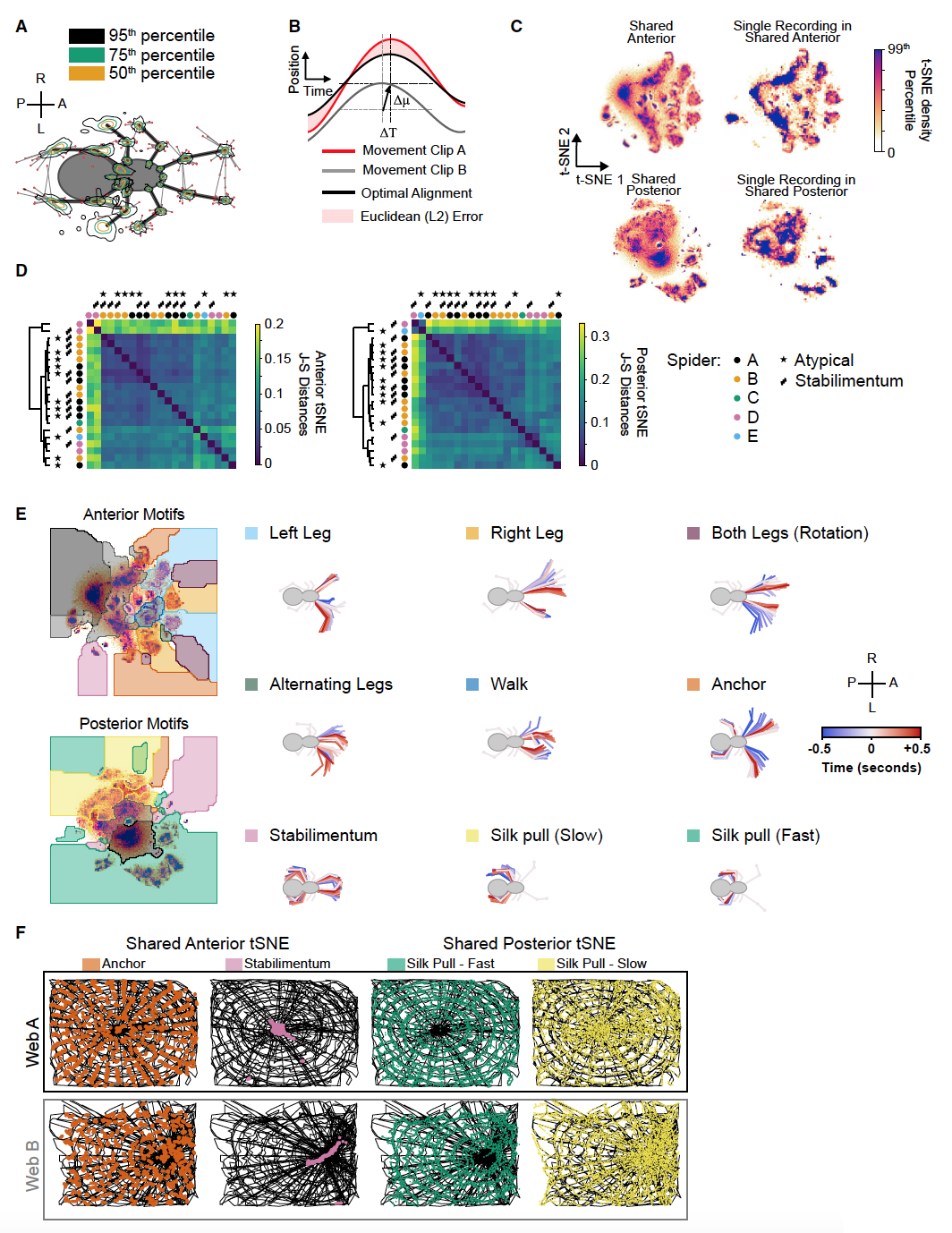
Изображение №3
Чтобы исследовать поведение паука в более точной пространственно-временной шкале, ученые отследили динамику 26 точек на его теле (3А). Для этого 10000 кадров были вручную аннотированы и использованы для захвата двух сверточных нейронных сетей (CNN от convolutional neural networks), отслеживающих конечности — LEAP и DeepLabCut.
Поскольку поведение — это не просто статические позы конечностей, а движение этих поз во времени, ученые изначально попытались зафиксировать это движение, передав вейвлет-преобразование по координатам. Этот подход определяет каждый момент времени как вектор частот движения для каждой отслеживаемой координаты, в результате чего получается многомерное пространство возможных частот конечностей.
Однако было обнаружено, что часто нерегулярное и менее синусоидальное поведение пауков приводит к появлению зашумленных вейвлет-профилей, в отличие от повторяющихся движений. В качестве альтернативного подхода было решено определять каждый кадр интервалом в 880 мс с центром в этом кадре (3B). Этот метод хоть и ограничивал временное разрешение каждого кадра до 880 мс, зато позволял более надежно сравнивать непериодическое и нерегулярное поведение, которое плохо определялось бы с помощью вейвлет-подхода.
И выбранный метод, и вейвлет-метод определяют каждую точку времени как вектор всех отслеживаемых координат во временном окне с центром в этой точке. Если одно и то же поведение многократно повторяется в разные моменты времени, то эти моменты времени должны находиться рядом друг с другом в многомерном пространстве. Чтобы сделать это пространство более интуитивно понятным, его можно спроецировать в два измерения, используя стохастическое вложение соседей с t-распределением (t-SNE). Чтобы сделать возможными перекрестные сравнения записей, общее пространство t-SNE было построено из 200000 временных окон, построенных равномерно из 21 записи. Все моменты времени записи впоследствии были вложены* в это общее пространство без изменения его фиксированной структуры.
Вложение* — специального вида отображение одного экземпляра некоторой математической структуры во второй экземпляр такого же типа.
Поскольку движения передних и задних лапок часто выполняются независимо, передние и задние лапки были вложены отдельно (3С, 3D), что улучшает распознавание движений. Для количественной оценки сходства вложений t-SNE использовалась дивергенция Дженсена-Шеннона (J-S от Jensen-Shannon), поскольку она не полагается на предварительные предположения о геометрии плотности (3D). Чтобы лучше различать поведение, определяемое t-SNE, к плотностям t-SNE была применена функция преобразования, чтобы разделить их на дискретные области.
Некоторые из поведений в этих областях можно легко аннотировать как отдельные поведения, которые наблюдались ранее (взмахи лапками, закрепление шелка, построение маскировки и вытягивание шелка), подтверждая полезность этого автоматизированного подхода (3E).
Видео №3: репрезентативные примеры 1-секундного движения для каждого кластера движений (соответствует изображению №3).
Видео №4: произвольно отобранные примеры 1-секундного движения для каждого кластера движений, иллюстрирующие стереотипные и атипичные движения (соответствует изображению №3).
Судя по данным, передние лапки по большей степени использовались для исследования, а потому совершали больше всего движений. Поскольку паук строит паутину в полной темноте, частые движения передних лап нужны для поиска шелковых нитей (3E). Передние лапы также участвовали в передвижении паука. Поскольку на передние маркеры (коих было 26) влияла трехмерная ориентация просомы (передняя доля тела паука), траектории передних конечностей также сигнализировали о вращении, закреплении шелка и создании маскировки.
Задние лапки в основном использовались для вытягивания и/или направления шелка из брюшка. Даже ходьба по шелку в основном выполнялась медиальными и передними лапками.
Наблюдалось четыре основных тактики вытягивания шелка — две относились к быстрым движениям и две — к медленным (отмечено желтым и зеленым на 3E). Более медленное вытягивание шелка доминировало на этапах строительства прото-паутины, радиусов и вспомогательной спирали. Этот вариант вытягивания происходил при достаточно высоком значении частот.
Более быстрое вытягивание относилось к процессу «расчесывания» захватывающего шелка (к которому, прилипает добыча). Частота лапок в этом случае была постоянной (9 Гц).
Малозаметное натягивание шелка, характерное для стадии создания маскировки, аналогичным образом можно определить по задним лапкам (отмечено розовым на 3E).
Иногда поведение закрепления шелка можно было определить по задним лапам, но траектории задних конечностей были малозаметны во время сгибания брюшка по сравнению с траекториями передних конечностей из-за разницы в их системе координат (отмечено оранжевым на 3E).
Из-за регулярной геометрии паутины некоторые их вышеперечисленных поведений происходили с регулярными интервалами у всех особей.
Фиксация шелка, к примеру, регулярно происходила, когда паук прикреплял захватывающий шелк к радиусам. Поэтому это часто происходило в местах, где паук пересекал радиальные линии (3F). Поведение, связанное с маскировкой, также наблюдалось исключительно в той области паутины, где паук ее создавал.
Полученный набор данных, описывающих разные типы поведения на разных этапах строительства паутины, позволяет автоматизировать процесс идентификации этих самых этапов без необходимости в ручном аннотировании каждого кадра записи наблюдений.
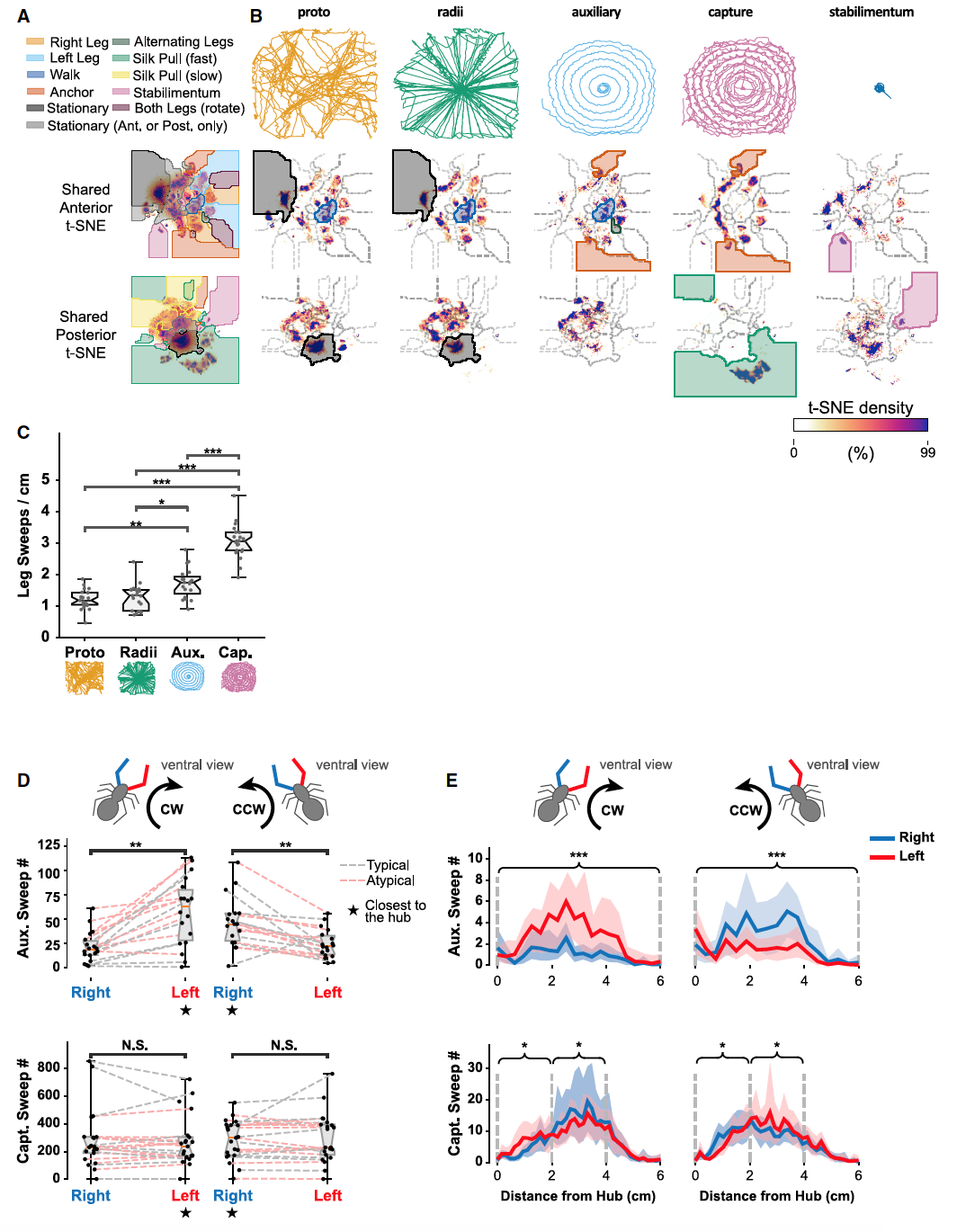
Изображение №4
Поскольку построение различных элементов сети, таких как радиусы и спираль захвата, разделено как в пространстве, так и во времени (1F), поведение, которое в основном происходило в определенных областях сети (3F), также демонстрировало сильные временные отклонения. Это стало более очевидным, когда плотности поведения в t-SNE пространстве были разделены на этапы построения сети (4A и 4B).
Движения передних конечностей, взятые во время стадий строительства прото-паутины и радиусов, были очень похожими, как и ожидалось, из-за схожей конструкции этих элементов паутины. Оба этапа характеризовались строительством длинных линий, охватывающих большую часть пространства. Следовательно, на этих этапах было больше всего ходьбы, чем на других (отмечено синим на 4B). Также было больше и пауз, что определяется динамикой передних или задних конечностей (отмечено серым на 4B). К примеру, 90% этапа прото-паутины занимали именно паузы (1I). Построение радиусов также включало длительные паузы, хотя их продолжительность была не такой большой. Во время строительства вспомогательной спирали наблюдаемая картина поведения была похожа, но пауз было меньше, а их продолжительность короче. Однако на этом этапе усиливалось поведение, связанное с закреплением шелка, так как вспомогательная спираль неоднократно пересекает радиусы, к которым прикрепляется. Закрепление шелка присутствовало и во время строительства спирали захвата, где также было мало паузы небольшой продолжительности (4B и 1I). Также было мало ходьбы, так как паук в основном сосредоточен на прикреплении спирали захвата, которая ортогональна радиусам (отмечено синим на 4B). Частое бездействие передних лапок не являлось неподвижностью (отмечено серым на 4В), так как в этот момент задние лапки занимались расчесыванием нитей захвата. Это поведение было самым ярким в рамках данного этапа строительства, так как большую часть времени задние конечности занимались именно расчесыванием (отмечена зеленым на 4B). Если паук создавал маскировку, то движения передних и задних лап было ритмичным и уникальным для данного этапа (отмечено розовым на 4B). Взмахи лапками происходили на всех этапах строительства паутины (светло-оранжевые и светло-синие области на 4A), но сильнее были выражены во время создания вспомогательной спирали и спирали захвата (4C).
Паук часто менял направление вращения во время создания спиралей. Во время строительства вспомогательной спирали наблюдалось явное смещение для взмахов лапки, ближайшей к ступице, независимо от угловой траектории (4D). На этом этапе паук разворачивается по спирали наружу от центра, поэтому ранее собранная вспомогательная спираль находится ближе к ступице.
Изменения в активности разных лапок (в аспекте опоры) отражает изменения в структуре строящейся паутины, а также изменение целей паука по мере того, как он переходит от одного этапа к другому.

Изображение №5
Несмотря на то, что разные поведения (например, взмахи лапками) были общими на разных этапах построения сети, то, как они координировались, необязательно было одинаковым. В частности, спираль захвата была наиболее регулярной. Задние лапы в основном выполняли расчесывание шелка с последующими регулярными этапами закрепления захватывающих нитей по радиусам (5B).
Видео №5: пример траектории спирали захвата, показывающий время t-SNE кластера движения задних и передних лап (соответствует изображениям №3 и №5).
Передние ноги либо оставались неподвижными, либо выполняли взмахивающие движения. Регулярное развитие этого поведения можно увидеть на графике вероятности их появления относительно поведения закрепления шелка (5C).
После прикрепления нитей захвата к радиусам процесс расчесывания начинается достаточно медленно (желтые квадраты на 5C), за которым следуют взмахи лапок (оранжевый и желтый квадраты на 5C), короткие паузы (черные квадраты на 5C), а затем быстрое расчесывание (зеленые квадраты на 5C). Регулярность этих поведенческих переходов отражает регулярность спирали захвата и координацию, которая необходима для обеспечения правильного построения этой важной части паутины.
Несмотря на то, что быстрое расчесывание шелка является уникальным поведением для этапа строительства спирали захвата, другие действия (взмахи лап или закрепление нитей) должны координироваться по-разному во время других этапов, так как потребности в строительстве различаются на каждом этапе.
Поскольку пауки не используют свое зрение для построения паутины, вытягивание шелка часто сочеталось с движениями передних лап (5D). Переходы от [взмахи лап + натяжение шелка] к [неподвижные передние лапы + натяжение шелка] отражают то, что паук сначала исследует местную среду своими передними лапами, определяет линии, а затем цепляется за них, в то время как задние лапы продолжают вытягивать шелк.
Определенные переходы были более характерны для определенных этапов и отражали более частое возникновение исходного состояния. Например, переходы в/из поведения ходьбы и поворотов более вероятны в состояниях прото-паутины и радиусов, что отражает важную роль, которую это поведение играет при сборке конкретно этих частей паутины.
Видео №6: пример радиальной траектории, показывающий время t-SNE кластера движения задних и передних лап (соответствует изображениям №3 и №5).
Несмотря на то, что большинство поведений являются общими для разных этапов, различия в относительной выборке поведений и их переходы подразумевают, что каждая стадия построения сети отражается не только в траектории центроида паука, но и в его поведении во время перемещения по паутине.
Изображение №6
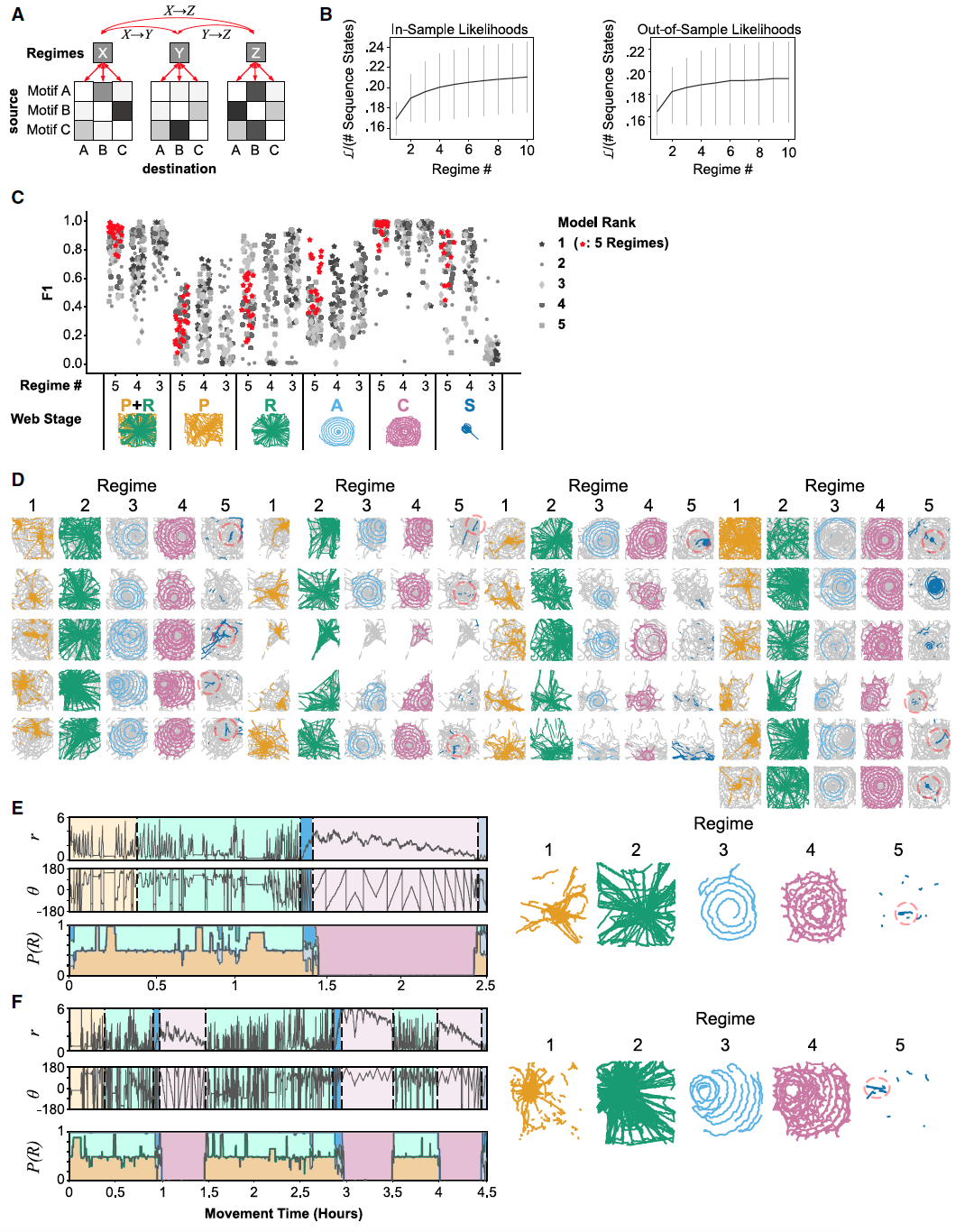
Проанализировав данные наблюдений, ученые пришли к выводу, что смогут предсказывать этап строительства паутины только на основе поведения паука. Была создана иерархическая скрытая марковская модель (HHMM от hierarchical hidden Markov model) со скрытыми родительскими состояниями верхнего уровня и нескрытыми дочерними состояниями, представляющими различные мотивы движения (6A).
Родительские состояния представляли (скрытые) режимы с различными лежащими в основе поведенческими переходными вероятностями, тогда как дочерние состояния представляли сами поведенческие переходы. Модели неоднократно обучались на подмножестве данных (в выборке) и использовались для прогнозирования изменений режима в данных вне выборки (6B).
Как и ожидалось, точность моделей увеличивалась с увеличением количества режимов. Но уже после третьего режима точность быстро стабилизировалась (6B). Для каждой модели были рассчитаны баллы F1, чтобы оценить, насколько хорошо эти модели соответствуют аннотированным вручную этапам строительства паутины.
В результате моделирования было определено пять режимов, соответствующих пяти этапам строительства. При этом модель смогла определить этапы вспомогательной спирали, маскировки и спирали захвата (6C). Различия между этапами прото-паутины и радиусов были смешанными, с преобладанием первого в аспекте количества пауз (6E). Это предсказание полностью соответствует наблюдениям. Другими словами, HHMM было так же сложно полноценно различить эти два этапа, как и наблюдателю. Однако, как комбинированное состояние, эти два режима точно соответствовали этапам прото-паутины и радиусов (6C—6E). Модель также успешно идентифицировала этап создания маскировки (6D).
Переходы между режимами точно соответствовали переходам между этапами, определяемым траекторией паука, даже для особей, у которых было нетипичное прогрессирование строительства (6F).
В совокупности эти данные говорят о том, что созданная модель может вполне успешно определять тот или иной этап строительства паутины, основываясь лишь на данных о поведении паука.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня труде ученые смогли детально описать каждый этап строительства паутины, а также охарактеризовать поведение паука в соответствии с каждым его действием.
Как заявляют сами авторы исследования, дабы понять, как мозг столь маленького существа работает, необходимо детально задокументировать все, что оно делает в процессе создания столь сложной структуры как паутина.
В ходе опытов для пауков была подготовлена специальная камера, где они могли плести свои сети в полной темноте, как они и любят. Записи процесса анализировали сначала вручную, что занимает уйму времени и не всегда может дать стопроцентно точный результат. А все потому, что человеку приходиться отслеживать положение всех лапок, коих у паука предостаточно, в каждом кадре видеозаписи. Посему на смену людям пришла машина, а точнее программа, обученная отслеживать положение конечностей тела паука кадр за кадром. В результате удалось создать детальную картину происходящего с точки зрения динамики нитей и самого паука.
Оказалось, что пауки весьма предсказуемы в своих действиях. Правильнее сказать, что строят они свои сети по очень точным стандартам, которых придерживаются все особи одного вида и даже особи других видов. Весь процесс строительства разделяется на определенные этапы, на каждом из которых паук совершает определенный набор действий, результатом которых будет завершение той или иной части сети (каркас, радиусы, дополнительная спирать, спираль захвата и т.д.).
Учитывая столь точный план стройки, ученые предположили, что это можно предсказать. Использовав моделирование ученые смогли предсказать, на каком этапе строительства находится паутина исключительно по положению лапок паука. Окончательный результат паучьих трудов может слегка отличаться между особями, но схема остается прежней, а потому модель предсказания отлично работает с разными паутинами.
В будущем ученые намерены провести опыты с препаратами, меняющими сознание, дабы понять, какие именно области мозга паука отвечают за какие именно этапы строительства.
По словам ученых, их труд позволяет не только больше узнать о пауках, но и создает фундамент для лучшего понимания нас самих. Ведь понять, как работает столь сложная система, как мозг человека, будет гораздо проще, если для начала мы поймем как работает мозг хотя бы паука.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


