
С чем у вас ассоциируется лето? Для кого-то это период долгожданного отпуска, для кого-то — каникул, а для кого-то — жара, духота и дискомфорт. Если же рассматривать лето с точки зрения гастрономии, то это период овощей, фруктов и ягод, которые мы любим не только за их вкус и пользу, но и за внешний вид. Как мы знаем из начального курса биологии, плоды многих растений обладают теми или иными свойствами, целью которых является привлечь потенциального гурмана. Это важная составляющая тактики расширения ареала произрастания. Подавляющее большинство плодов имеют яркий и сочный цвет, оповещающий об их вкусности. Главным источником того или иного окраса у ягод являются пигменты в кожуре, однако это не единственная методика окрашивания. Ученые из Бристольского университета выяснили, что калина лавролистная (Viburnum tinus) использует липидные наноструктуры в клеточных стенках для окрашивания своих ягод, что является ранее неизведанным вариантом структурной окраски. Что такого необычного в этих липидных наноструктурах, за счет чего они придают ягодам темно-синий окрас, и какое практическое применение сего открытия? Свет на эти вопросы прольет доклад ученых. Поехали.
Результаты исследования
Главным героем данного труда является калина лавролистная (Viburnum tinus / калина тинус) — вечнозеленый куст или дерево до 6 метров в высоту, произрастающее в Средиземноморском регионе.

Viburnum tinus во время цветения.
В течение года калина тинус несколько раз плодоносит темно-синими ягодами, обеспечивая пищей множество видов птиц, среди которых черноголовая славка (Sylvia atricapilla) и зарянка (Erithacus rubecula). Как и для любых других ягодных растений, птицы для калины V. tinus являются основным методом распространения семена на новые территории.

Sylvia atricapilla (слева) и Erithacus rubecula (справа).
На первый взгляд в этом растении нет ничего особенного. Красивый вечнозеленый кустарник, радующий эстетов среди людей и гурманов среди птиц. Однако детальное рассмотрение ягод говорит об обратном. Пока другие растения используют различные химические соединения для окрашивания своих плодов, калина тинус использует структурное окрашивание. Хотя ранее считалось, что синий цвет ягод V. tinus вызван присутствием антоциановых пигментов в их кожуре, что, естественно, не является истиной.
Структурные цвета достаточно распространены среди представителей фауны: крылья бабочек, панцири жуков, перья павлинов и т.д. Цвет в их случае образуется за счет наноразмерных структурных особенностей поверхности, вызывающих интерференцию видимого света.

Примеры структурных цветов в природе: А — гибискус тройчатый (Hibiscus trionum); В — жук тамамуси (Chrysochroa fulgidissima); С — бабочка вида Morpho rhetenor; D — комар обыкновенный (Culex pipiens); Е — морская мышь (Aphrodita aculeata); F — жук вида Pachyrhynchus argus; G — бабочка вида Parides sesostris.
Особенность калины тинус, привлекшая внимание ученых, заключается в том, что она не только демонстрирует новый механизм структурного окрашивания, но и является одним из немногих растений, способных на это.
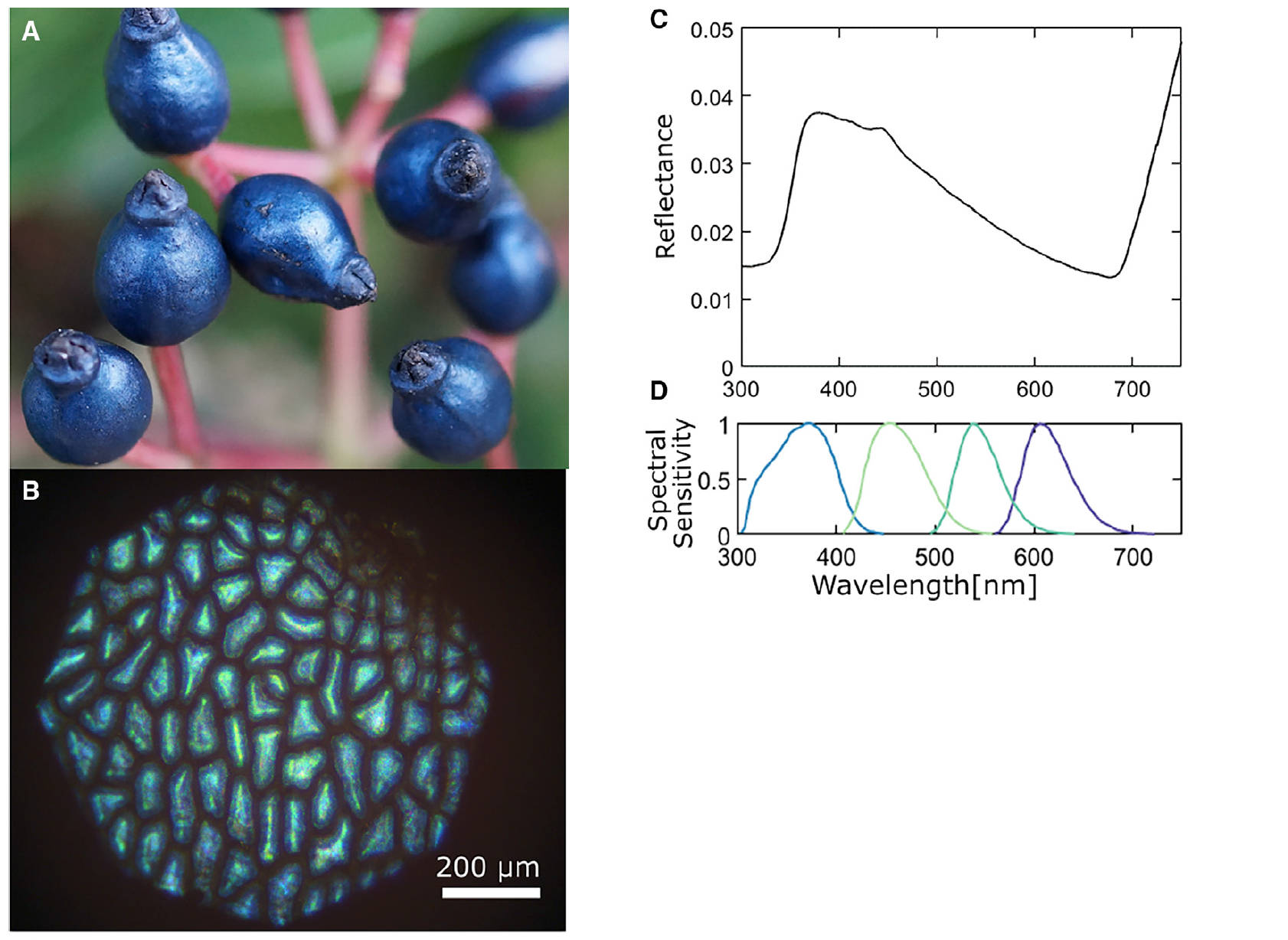
Изображение №1
Плоды V. tinus (1C) отражают свет направленно (придавая ему металлический вид) в синей и ультрафиолетовой области спектра. Поляризация отраженного света в основном сохраняется, что указывает на то, что окраска является структурной, а не пигментной, возникающей в результате отражения от высокоструктурированной клеточной стенки внешнего эпикарпа* (2А).
Эпикарп* — внешний слой плода.
При диссекции этой ткани выделяется темно-красный антоциановый пигмент. Свет, который не отражается фотонной структурой, поглощается темно-красным пигментом под ним (2А и 3C).
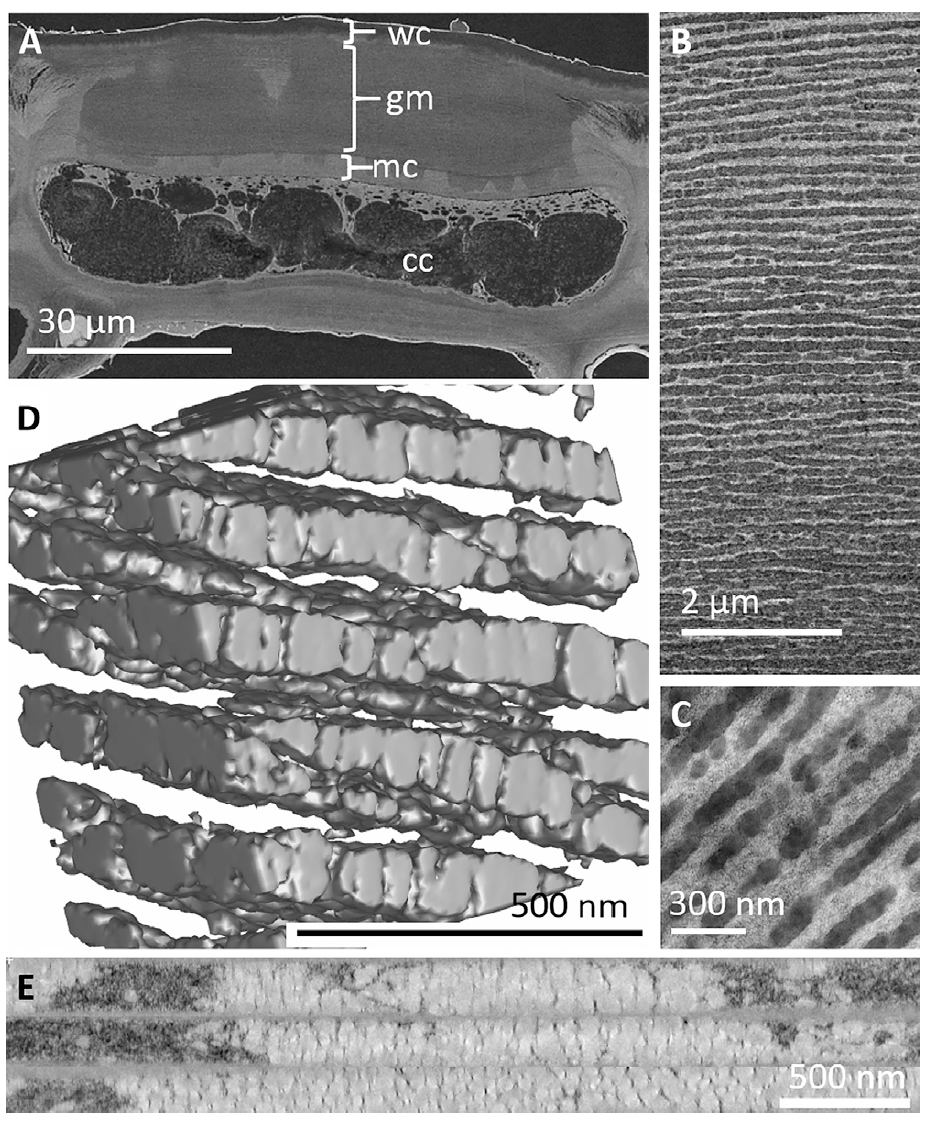
Изображение №2
Данное поглощение предотвращает обратное рассеяние света, увеличивая заметность синего отражения от внешней стенки клетки и, таким образом, визуально улучшая синий окрас.
Из этих наблюдений уже можно сделать вывод, что цвет плодов V. tinus является результатом комбинации физической наноструктуры, которая избирательно отражает волны синего света, и базового слоя пигментов, усиливающих синий цвет. Другими словами, сопряжение химии и физики.
Чтобы охарактеризовать наноструктуры, создающие синий цвет в плодах V. tinus, ученые использовали несколько методов электронной микроскопии.
Сканирующая электронная микроскопия свежих тканей (2А) четко показывает наличие толстой (10–30 мкм) многослойной структуры, параллельной поверхности плода и встроенной в клеточную стенку самых внешних эпикарпальных клеток. Поверхность плода покрыта восковой кутикулой (2–6 мкм) поверх слоистой структуры. Слоистая архитектура занимает большую часть внешней клеточной стенки в области между кутикулой и богатой на целлюлозу первичной клеточной стенкой. Слои имеют толщину от 30 до 200 нм и покрывают всю клетку.
Просвечивающая электронная микроскопия показывает, что эта архитектура состоит из множества слоев мелких пузырьков, которые отличаются от матрицы по способности рассеяния электронов и показателю преломления.

Изображение №3
Снимки, полученные в ходе сканирующей и просвечивающей микроскопии, показывают, что матрица, по-видимому, содержит ключевые компоненты типичных стенок растительных клеток, а именно целлюлозу, гемицеллюлозу и пектин. Окрашивание рутениевым красным (3D) показывает значительное содержание пектина, а электронограмма демонстрирует присутствие целлюлозы по характерным дифракционным кольцам кристалла природной целлюлозы.
Стоит отметить, что контрастные слои являются дискретными и остаются отличными друг от друга, но значительный беспорядок вносится непараллельными соседними слоями и неравномерностью их глобулярной структуры.
Томография стенки эпидермальных клеток (2E) показывает, что эти глобулярные везикулы организованы в объединенные слои, через которые матрица клеточной стенки целлюлозы остается соединенной мостиками и нитями (2B и видео ниже).
Модель глобулярной многослойной структуры (соответствует изображению 2D).
Из этого следует, что глобулярная многослойная структура эпидермиса плодов V. tinus состоит из липидов, встроенных в матрикс клеточной стенки, с использованием различных методов.
Далее ультратонкие срезы эпидермиса плодов подвергались воздействию хлороформа. Этот анализ весьма показателен, так как растворимость в неполярных органических растворителях является явным признаком присутствия липидов.
ПЭМ-снимки одного и того же участка образца до (3A) и после (3B) воздействия хлороформа показывают, что глобулярная структура была удалена обработкой. На последнем изображении контраст многослойной глобулярной фазы снижен, а пустые структуры внутри матрицы остаются видимыми. Для сравнения, воздействие воды не изменило ультраструктуру или контраст изображения глобулярного многослойного материала, указывая на то, что материал можно экстрагировать только неполярными растворителями. Кроме того, когда в процессе химической фиксации ученые использовали забуференный имидазолом тетраоксид осмия (C3H4N2/OsO4), который связывается с липидами, глобулярные слои окрашивались, что подтверждает их липидную природу.
А когда использовали рутениевый красный, который связывается с пектином, матрица клеточной стенки окрашивалась, в то время как глобулярная структура удалялась ввиду отсутствия имидазольного буфера.
Во время всех вариаций окрашивания, примененных во время исследования, наблюдались темные очертания вокруг глобул (3Е). По мнению ученых, это может указывать на присутствие липидной мембраны, теоретически необходимой на интерфейсе между гидрофобными молекулами и гидрофильными полисахаридами вторичной клеточной стенки.
Ученые напоминают нам, что липиды состоят из разнообразных молекулярных структур, обычно классифицируемых как воски, жиры и масла, в зависимости от их температуры плавления.
На поверхности эпидермиса растений можно легко найти воски, образующие водостойкую восковую кутикулу. Воски также включают в себя множество молекулярных структур, но преобладающим компонентом остаются алканы, которые фактически не перевариваются, т.е. не имеют питательной ценности для птиц. А вот жиры и масла, напротив, являются жизненно важными пищевыми компонентом, поскольку содержат гораздо больше энергии на единицу объема, чем в крахмале или в белках. Жиры обычно можно найти в большом объеме в семенах, т.е. глубоко внутри плода.
В случае плодов V. tinus непосредственная близость глобулярной структуры как к крупным, богатым энергией семенам, так и к восковидной внешней кутикуле делает различие между воском и жирами особенно важным для понимания функционального значения и происхождение данной структуры. Следовательно, необходимо определить, являются ли липидные глобулы неперевариваемым воском или питательными жирами и маслами. Для этого была использована световая микроскопия.
Срезы тканей плода V. tinus инкубировали с нильским синим A (пигмент), который окрашивает богатую глобулами область клеточной стенки V. tinus в синий или сине-фиолетовый цвет (3C). А это говорит о том, что глобулы представляют собой свободные жирные кислоты, а не полимер кутина (компонент кутикулярной мембраны), который окрасился бы в розовый или красный цвет.
Кроме того, электронограмма многослойной глобулярной структуры показывает четкий кольцевой узор, отличающийся от диаграммы клеточной стенки целлюлозы с характерными двумя кольцами из кристаллов целлюлозы. Этот образец указывает на то, что липидные тела, вероятно, являются кристаллическими и, следовательно, являются гомогенными мономерными липидами, а не полимеризованными молекулами, такими как кутин.
Чтобы подтвердить, что наблюдаемая смешанная структура, состоящая из целлюлозной матрицы и слоистых липидных глобул, ответственна за синюю отражательную способность плодов V. tinus, ученые смоделировали ее оптический отклик. Для этого ученые исследовали две математические модели: двумерный массив сфер и усреднение по множеству одномерных двухфазных мультислоев.
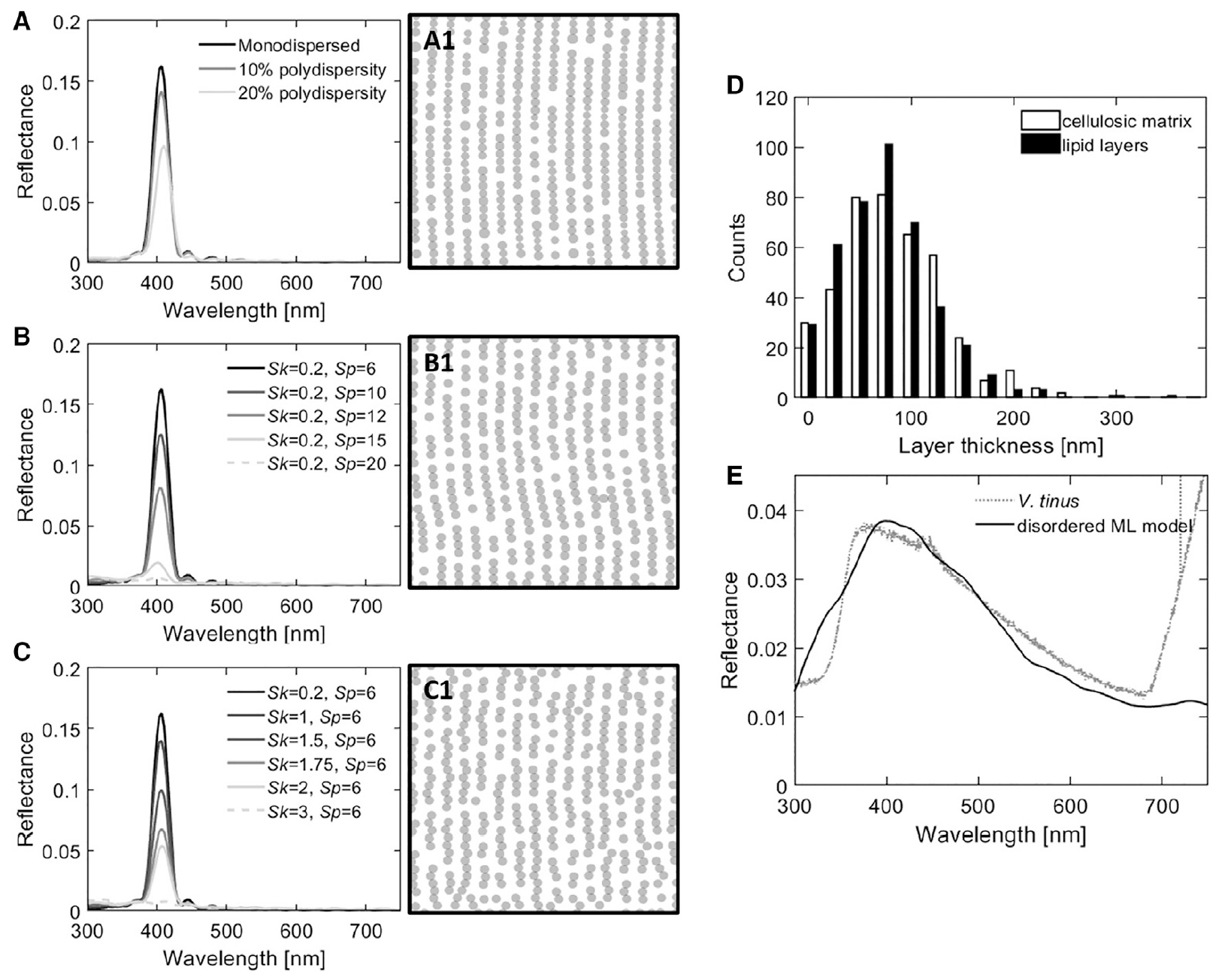
Изображение №4
Алгоритм обратного проектирования был использован для моделирования структуры в двумерном пространстве в виде серии глобулярных скоплений. Схемы на 4A—4C соответствуют смежному смоделированному спектру отражения. Этот алгоритм позволяет независимо вводить различные типы беспорядка в глобулярный мультислой, настраивая размер и структуру, т.е. преобразование Фурье положения частиц.
В процессе моделирования были изучены: оптический отклик слоистых липидных глобул с разной степенью вариации диаметра глобул (4A); беспорядок в углах между соседними глобулами (параметр Sp, 4B) и беспорядок на среднем расстоянии между соседними глобулами (параметр Sk, 4C).
Введение различных типов беспорядка (4A—4C) всегда оказывало одинаковый эффект на оптический отклик глобулярного мультислоя, а именно, снижение пиковой интенсивности.
Таким образом, вместо того, чтобы рассматривать каждый элемент беспорядка по отдельности, структура и материальный состав клеточной стенки V. tinus были аппроксимированы неупорядоченными одномерными мультислоями с показателями преломления, соответствующими целлюлозе (n = 1.55) и типичному растительному липиду (n = 1.47). Распределение толщины обоих материалов показано на 4D. А отражательная способность, смоделированная с использованием средних значений по одномерным слоям, показана на рисунке 4E.
Введение беспорядка, наблюдаемого в измерениях поперечного сечения, в модель когерентного упорядоченного отражателя расширяет его спектр отражения.
Если модель позволила ученым понять, как именно ягоды V. tinus получают свой окрас, то вопрос касательно надобности такого механизма моделирование не охватывает.
Наибольшее межвидовое взаимодействие у V. tinus связано с птицами, питающими ягодами этого удивительного растения. Сравнение со спектральной чувствительностью синицы (1D) показывает, что цвет ягод находится в пределах визуально значимого для птиц данного вида диапазона.
Ягоды, естественно не парят в воздухе, а прикреплены к веточкам, на которых растут листья — визуальный фон. По большей степени, фон зеленого цвета, ввиду доминирующего пигмента хлорофилла в листьях. Хлорофилл имеет широкую спектральную характеристику с пиком при 550 нм и незначительной отражательной способностью ниже 500 нм, благодаря чему цвет плодов V. tinus хроматически контрастирует с листвой. Другими словами, на фоне таких листьев ягоды выглядят еще заметнее.
Учитывая, что зрительные сигналы для птиц часто являются приоритетными, липидная структурная окраска ягод V. tinus может служить сильным визуальным сигналом для голодных птиц.
Если же учесть, что цвет пищи для птиц может быть первичным параметром съедобность, то цвет ягод V. tinus сигнализирует о том, что они съедобны и питательны.
Связь цвета плода и его питательной ценности исследовалась и ранее. По некоторым данным темные плоды растений из Бразильского региона богаты углеводами, а темные плоды растений Средиземноморья богаты липидами.
Ученые считают, что в случае V. tinus синий цвет является сигналом о том, что в ягодах много питательных липидов, которые, к слову, и создают этот окрас.
Такой метод передачи сигналов ученые называют «честным» или «прямым», когда контекст сигнала соответствует его источнику (синий цвет из-за липидов — высокое содержание липидов). Такой метод сигнализации достаточно затратен, ибо использование классической пигментации было бы проще для растения. Тем не менее, отдача, которую получает V. tinus в виде привлечения внимания птиц разных видов, судя по всему, перекрывает этот недостаток.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
Цвет является важной составляющей визуальной информации, которую живые организмы получают об окружающем их мире. Многие животные используют свой окрас для маскировки, привлечения партнеров или для отпугивания неприятелей. Среди растений также присутствуют некоторые их этих тактик, но самой значимой является поддержание межвидовой связи. В случае V. tinus птицы являются основными партнерами этого растения, необходимыми для распространения семян на большие расстояния, что значительно увеличивает ареал произрастания V. tinus и, следовательно, шансы на выживание вида.
Вкусовые качества плодов многих растений зависят от того, насколько они хотят привлекать к себе внимание животных тех или иных видов. Некоторые плоды будут вкусными, например, для конкретных видов птиц, тогда как для всех остальных они будут практически несъедобны. В такой сложной системе как межвидовая связь важную роль играет степень коэволюции видов растений и животных, которые ее формируют.
Синий цвет калины лавролистной заключается в его нестандартном происхождении — наноструктуры липидов, содержащиеся в стенках эпидермальных клеток ягод V. tinus. Данная методика окрашивания (структурное), особенно за счет липидов, среди растений на данный момент обнаружена только у V. tinus. Помимо этого липидное окрашивание может говорить птицам о большом содержании липидов в ягодах, как бы странно это ни звучало.
Честные сигналы, происхождение которых соответствует их контексту, достаточно редки в природе. Объяснение этой редкости достаточно простое. Представьте себе, что у вас есть булочная. Вы хотите привлечь больше покупателей, а потому раздаете флаеры. Следовательно, сигнал имеет один контекст (у нас вкусные булочки), но его происхождение — другой (у меня нет булочки, а нечто получше — рисунок булочки, т.е. флаер это лишь клочок бумаги). Если же вы будете раздавать булочки, то это будет честное сигнализирование, но куда более затратное.
Ранее многослойные липидные архитектуры, как у ягод V. tinus, не были замечены в биоматериале. В прошлом не было столь развитых инструментов и методик, как сейчас, потому многие детали были зафиксированы неверно или вовсе были упущены.
В будущем ученые намерены провести анализ других растений, которые теоретически могут также обладать подобными липидными наноструктурами и, следовательно, нестандартным методом окрашивания плодов. Кроме того, ученые считают, что их исследования могут способствовать созданию более безопасных пищевых красителей.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


