
Лед холодный, огонь горячий, а кактус лучше не обнимать: как мозг формирует сенсорную память
На протяжении всей жизни наш мозг непрерывно собирает и обрабатывает информацию. Часть этой информации кодируется в виде долговременной памяти, а другая часть загружается в кратковременную. Это может быть что угодно: недавно прочитанная книга, фамилия новых соседей, день рождения тещи или напоминание вынести мусор. Но, как мы все прекрасно знаем из личного опыта, некоторая доля информации частенько забывается. Однако «выборочный склероз» крайне редко затрагивает сенсорную память. Сенсорная информация, которую мы получаем из окружающей среды, помогает нам с этой средой взаимодействовать. К примеру, капот машины, долго простоявшей под открытым небом в жаркий летний день, явно холодным не будет — это результат не только логического мышления, но и сенсорной памяти. Проще говоря, вода мокрая, огонь горячий, а кактус колючий, принцип остается тот же.
Однако, как именно наш мозг формирует память на основе сенсорных данных, пока неизвестно. Ученые из общества Макса Планка (Мюнхен, Германия) решили пролить свет на этот таинственный процесс. Какие участки мозга задействованы в процессе формирования сенсорной памяти, как это происходит, и как понимание этого аспекта поможет в расшифровке работы мозга в целом? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследований
Мы не живем в вакууме, вокруг нас постоянно что-то происходит. Дабы полноценно существовать в окружающей нас среде, нашему мозгу необходимо сбалансировано интегрировать внешние сигналы от сенсорных систем с внутренней информацией, отражающей накопленный опыт и знания.
Информация, обрабатываемая нашим мозгом, условно делится на два типа: нисходящая и восходящая. В первом случае речь идет о данных, получаемых из окружающей среды. Во втором — о накопленном опыте и имеющихся знаниях.
Слой неокортекса L1 получает ряд нисходящих входных сигналов из разных источников и, следовательно, может быть ключевым узлом для интеграции этих сигналов. Ученые отмечают, что одна заметная, но малоизученная проекция на L1 возникает из таламуса более высокого порядка. Ученые считают, что эти афференты* представляют собой источник зависимой от опыта нисходящей информации, отличной от классического восходящего кодирования, которое широко наблюдается в таламических ядрах первого порядка.
Афферент* — нейрон, передающий импульсы от рецепторов к головному или спинному мозгу.
В связи с этим предположением исследователи решили детально изучить функции таламо-кортикальной коммуникации в аспекте памяти. Основной акцент исследования был поставлен на афферентах слуховой коры, критической области ассоциативной памяти.
Слуховой таламус более высокого порядка (HO-MG от higher order medial geniculate, т.е. медиальное коленчатое тело высшего порядка) состоит из нескольких ядер, которые окружают слуховой таламус первого порядка (MGv от ventral division of the MGB, т.е. вентральный отдел медиального коленчатого ядра). Чтобы избирательно нацеливать экспрессию белка на HO-MG, избегая MGv, была использована условная экспрессия, опосредованная аденоассоциированным вирусным вектором (AAV), у мышей CR-IRES-Cre (1A и 1B).
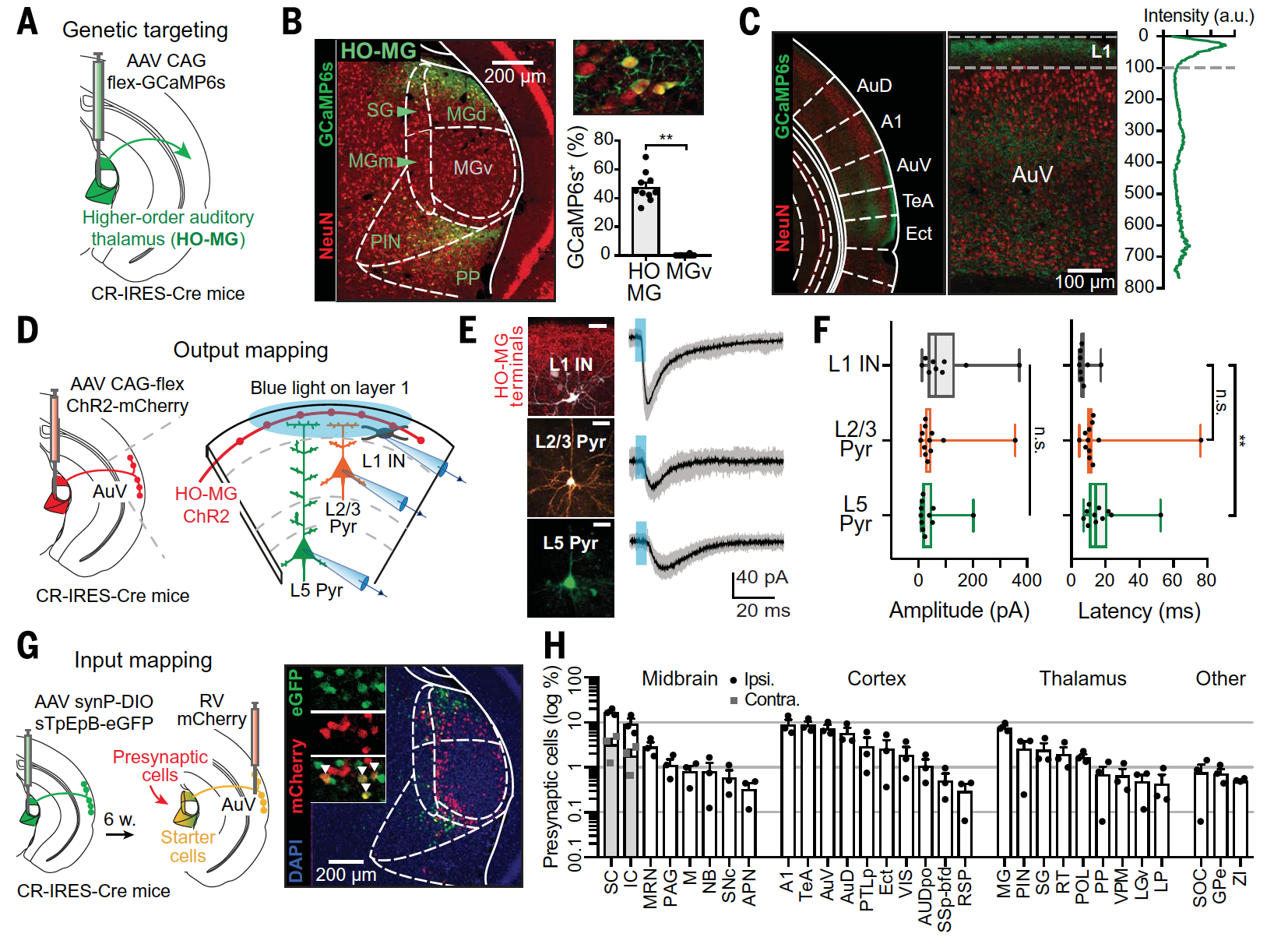
Изображение №1
Антероградное отслеживание* показало, что аксоны HO-MG демонстрируют средневисочный градиент плотности, который близко соответствует расположению областей коры, критических для ассоциативной памяти, с самой высокой плотностью в височных областях коры, включая вторичную слуховую кору (AuV) и временную ассоциациативную кору головного мозга (1С).
Антероградное отслеживание* — метод нейробиологического исследования, который используется для отслеживания проекций аксонов от их источника (тела клетки или сомы) до точки терминации (синапса).
Потому было решено сосредоточить дальнейшие усилия на изучении именно на височной области слуховой коры (AuV), где аксоны были сильно обогащены верхним L1 (1C).
Результаты исследования
Чтобы идентифицировать постсинаптические цели афферентов HO-MG в L1, было использовано картирование цепи с помощью канала родопсина-2 (ChR2) в срезах головного мозга при записи с интернейронов* L1 (L1 IN) и пирамидных нейронов* (Pyrs) в L2/3 и L5 (1D—1F).
Интернейрон* — нейрон, который связан исключительно с другими нейронами. Интернейрон противоположен, например, двигательным нейронам, которые иннервируют мышечные волокна.
Пирамидальный нейрон* — основной возбудительный нейрон мозга млекопитающих.
Световая стимуляция L1 приводила к возбуждению трех типов нейронов с одинаковой амплитудой (1E и 1F), но с разной вероятностью. В соответствии с синаптическими контактами, находящимися на дистальных дендритах Pyrs, токи возбуждения, зарегистрированные в L5, демонстрируют более длительные латентные периоды (1F).
Эти наблюдения показывают, что вход HO-MG L1 обеспечивает широкий контроль над активностью коры, которая может влиять на обработку цепи различными способами. К примеру, привлечение к процессу IN L1 может генерировать растормаживание нижних слоев, что особенно важно для ассоциативного обучения, а также может напрямую подавлять дистальные дендриты.
Вышеописанные результаты полностью согласуются с тем, что HO-MG играет роль основной нисходящей проекции. Предполагается, что нисходящие сигналы будут передавать информацию обратной связи из различных источников.
Далее ученым удалось идентифицировать сигналы, поступающие именно в нейроны HO-MG, проецирующие слуховую кору. Для этого был использован метод Cre-зависимого транссинаптического ретроградного отслеживания вируса бешенства (1G).
Данный эксперимент выявил различные источники сигналов (1H), включая некоторые области коры, промежуточные и глубокие слои верхнего двухолмия*, внешнюю капсулу нижнего двухолмия* и периакведуктальное серое вещество*.
Верхнее двухолмие* — одна из основных структур среднего мозга, слои которой отвечают за обработку визуальной информации (верхние слои) и за моторику (глубинные слои).
Нижнее двухолмие* — основное ядро среднего мозга слухового пути, которое принимает сигналы от нескольких периферических ядер мозгового ствола слухового пути, а также сигналы, полученные от слуховых центров коры больших полушарий.
Периакведуктальное серое вещество* — скопление серого вещества под водопроводом мозга, анатомически относящееся к покрышке среднего мозга.
Дабы точно определить, передают ли афференты HO-MG информацию, связанную с памятью, к коре головного мозга, ученые использовали дискриминативное обусловливание угрозы (DTC от discriminative threat conditioning), чтобы наделить слуховые стимулы зависимой от опыта релевантностью контролируемым образом. Обусловливание осуществлялось путем многократного сочетания одного из двух изначально нейтральных условных стимулов (CS от conditioned stimuli) с легкими ударами тока.
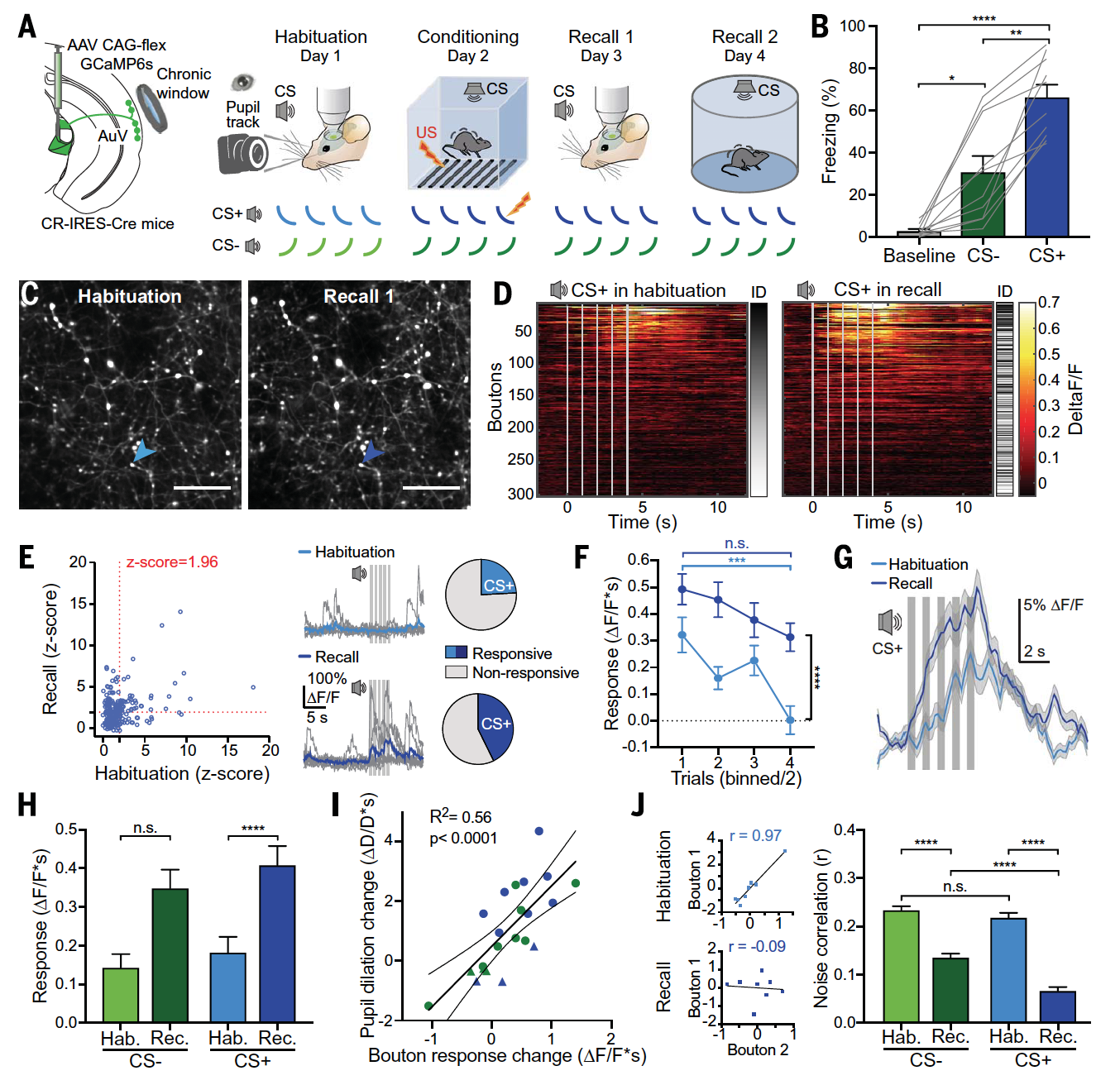
Изображение №2
Данный метод был объединен с двухфотонной кальциевой визуализацией бутонов* HO-MG, экспрессирующих GCaMP6, в L1 бодрствующих мышей с зафиксированной головой после идентификации AuV с помощью внутренней визуализации (2A).
Бутоны* — дистальные окончания телодендрий (ветвей) аксона.
Визуализация бутонов осуществлялась как после воздействия стимулов с ударом тока (CS+), так и после воздействия стимулов без тока (CS—) за день до фактических тестов и спустя день после них (2A).
Стимулы вызывали формирование памяти о возможной угрозе (удар током). Это можно было измерить за счет расширения зрачков во время тестов и за счет инстинктивного замирания мыши (которая уже участвовала в тестах) в камере, где ток не применялся (2А и 2В).
Для анализа были выбраны лишь те бутоны, что были идентифицированы на этапе привыкания (во время тестов с током) или вспоминания (в камерах без тока), что позволило провести более точный сравнительный анализ нейронной активности в ответ на стимулы (2С).
Стимуляция CS+ во время этапа привыкания у бодрствующих мышей с зафиксированной головой вызвала четкие ответы в подгруппе бутонов (2D и 2E). Однако синаптические ответы HO-MG быстро затухали до исходного уровня после нескольких испытаний (2F), указывая на то, что стимул, кодируемый афферентами HO-MG, очень чувствителен к новизне.
Другой отличительной особенностью была их заметно замедленная кинетика, при которой большая часть ответа происходила после каждого однократного стимула (2D и 2G), что сильно отличалось от ответов с короткой задержкой, обычно обнаруживаемых в MGv первого порядка.
А вот ответы на CS+ стимулы во время вспоминания продемонстрировали различные пластичные изменения, связанные с обучением, что приводит к гораздо более сильной активации бутонов HO-MG.
Во-первых, в то время как CS+ ответы отдельных бутонов либо увеличивались, либо уменьшались, общее количество ответных бутонов увеличивалось на 77% после обучения (2D и 2E). Во-вторых, наличие CS+ ответов было намного более устойчивым во время испытаний, чем на этапе привыкания (2F). В-третьих, CS+ ответы стали более привязанными по времени к началу стимуляции (2D и 2G) и предшествовали проявлению поведенческой памяти. И, в-четвертых, общий CS+ ответ сильно усиливался при обучении (2G и 2H), в основном во время стимуляции, но также, в меньшей степени, после ее отключения.
Изменения, наблюдаемые в ходе экспериментов, были значительно выше у мышей, тестируемых в условиях CS+ стимулов (с ударами током), чем у мышей, тестируемых в условиях CS— стимулов (2H).
Чтобы напрямую определить взаимосвязь между пластичностью и силой памяти, были измерены изменение зрачков мышей от этапа привыкания к этапу вспоминаниям.
Анализ изменений реакции бутонов показал, что индивидуальная вариабельность величины потенциации (синаптической передачи между двумя нейронами) сильно коррелировала с силой памяти и, таким образом, могла быть ее предшественником (2I).
Дополнительно была проанализирована корреляция активности между бутонами, которая может ограничивать содержание передачи информации. Корреляция шума от опыта к опыту между всеми парами бутонов показывала уменьшение активности на этапе вспоминания по сравнению с привыканием. Это было вызвано, в частности, уменьшением корреляции между бутонами, реагирующими на стимулы, и неотвечающими бутонами (2J), в то время как корреляция между реагирующими и невосприимчивыми бутонами сохранялась неизменной.
В совокупности эти данные демонстрируют, что HO-MG передают информацию о поведенческой значимости звуков в слуховую кору L1, которая зависит от предыдущего опыта как в краткосрочной перспективе (во время привыкания), так и в долгосрочной перспективе (после ассоциативного обусловливания).
Второй важный аспект, не зависящий от опыта, происходит из физических свойств сенсорных стимулов, таких как интенсивность звука. Чтобы выяснить, кодируют ли афференты HO-MG также и восходящую информацию, была проведена визуализация бутонов в ответ на стимулы в виде акустической последовательности с различной интенсивностью (3А и 3В).
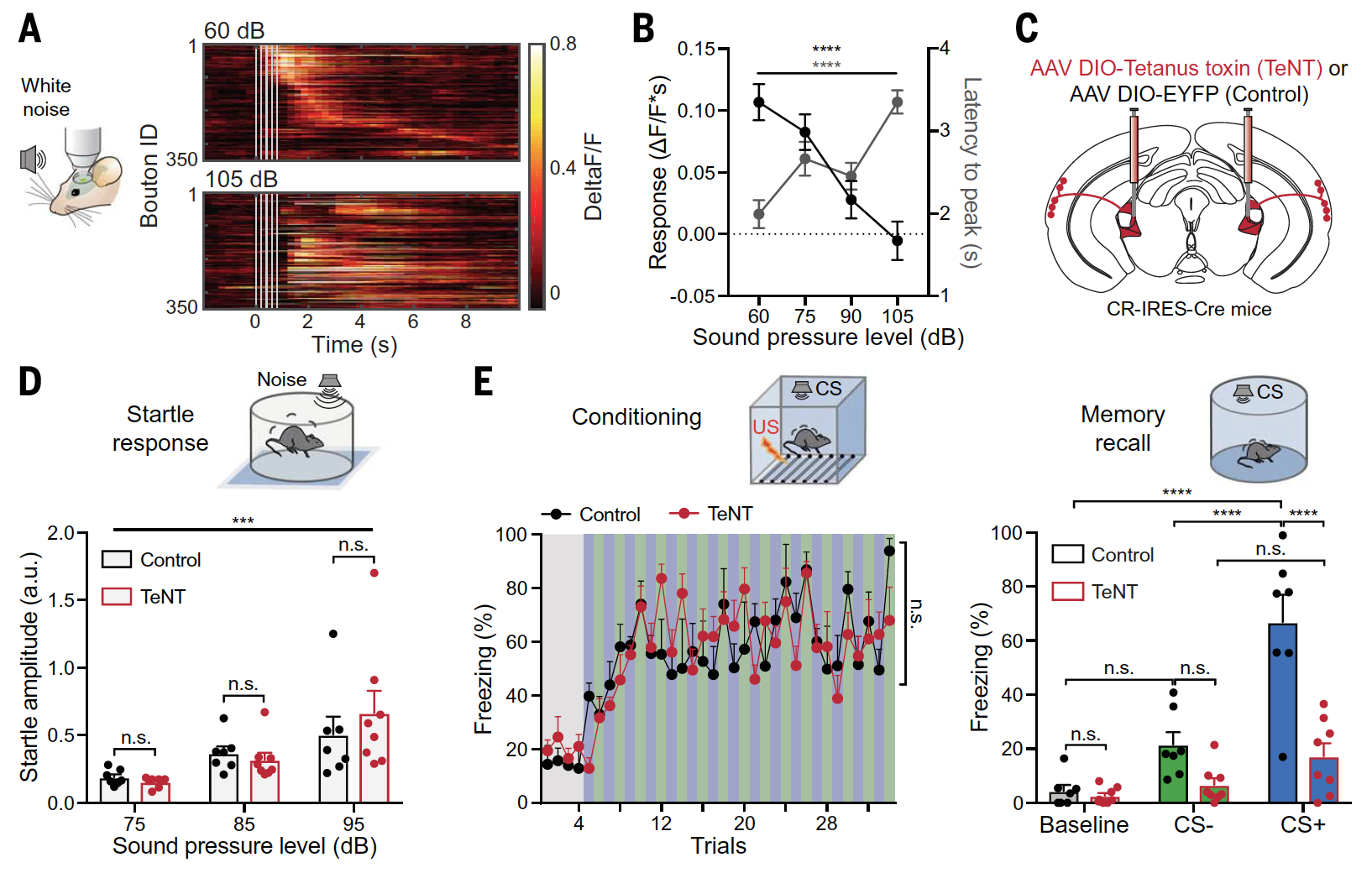
Изображение №3
В отличие от изменений, наблюдаемых после обучения, и типичного кодирования в таламических афферентах первого порядка, реакции бутонов во время акустической последовательности уменьшались с увеличением интенсивности стимула (3B).
Далее было проверено, что необходимо от сигналов HO-MG для формирования ассоциативной памяти. Для этого нейроны HO-MG были деактивированы с помощью тетанотоксина (TeNT), а базовая слуховая обработка осталась нетронутой (3С и 3D). Это привело к нарушению экспрессии памяти в случае CS+ стимулов наряду с дефицитом различения после дискриминативного обусловливания угрозы (справа на 3E).
При этом поведение во время обнаружения угрозы (удар током через пол тестовой камеры) и соответствующая реакция никак не изменились (слева на 3Е). Это указывает на то, что HO-MG критически необходим для ассоциативной слуховой памяти, но не является необходимым для базового восприятия угрозы и неслуховой памяти.
Принимая во внимание, что нисходящая информация обычно считается независимой от активности в локальной цепи, ученые предположили, что эти афференты могут находиться под функциональным контролем специализированного типа нейроглиаформного интернейрона, который плотно иннервирует L1 и экспрессирует белок NDNF (нейротрофический фактор, полученный из нейронов).
Поскольку эти нейроглиоформные клетки могут осуществлять опосредованную передачу посредством g-аминомасляной кислотой, они могут контролировать вероятность высвобождения бутонов HO-MG посредством пресинаптического ингибирования рецептора ГАМК типа B (4A).

Изображение №4
Сначала необходимо было понять, могут ли HO-MG в коре головного мозга подавляться посредством GABABR (ГАМК типа B).
Анализ срезов головного мозга показал, что агонист* GABABR баклофен снижает амплитуду постсинаптического тока (4B и 4C).
Агонист* — химическое соединение, которое при взаимодействии с рецептором изменяет его состояние.
Когда аксоны стимулировались серией световых импульсов (4D), баклофен усиливал передачу более высоких частот. Это указывает на то, что пресинаптическая активация GABABR может модулировать частотный состав афферентной информации.
Вышеописанные результаты идентифицируют HO-MG как сильно зависящий от опыта источник нисходящей информации, которая имеет решающее значение для ассоциативной памяти.
В L1 дендриты Pyrs являются центральным субстратом для интеграции и компартментализации различных афферентов.
Результаты исследования показывают, что эти механизмы будут вносить вклад в обработку сигналов от HO-MG. При этом была установлена ранее неизвестная форма модуляции афферентной информации, действующая на пресинаптические GABABR, которые могут динамически управлять синаптической передачей.
Зависимость от локального ингибирования не только означает, что информация, кодируемая бутонами дальнего действия, может отличаться от информации, кодируемой на уровне сомы, но также и то, что коллатерали (боковые или обходные пути) от отдельного аксона могут передавать различную информацию в различные целевые области мозга. Учитывая, что пресинаптические GABABR присутствуют в большом количестве синапсов, вероятно, что дополнительные афференты к кортикальному L1 подвержены этой форме модуляции.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Информация, обрабатываемая мозгом, может быть категоризована на два типа: нисходящая и восходящая. Связь между этими типами информации позволяет организму соответственно реагировать на внешний стимул.
В данном труде ученым удалось измерить реакцию отдельных таламических синапсов в неокортексе мышей до и после обучения (ряда тестов с определенными внешними стимулами). Анализ результатов показал, что нейтральные стимулы кодировались небольшими и временными реакциями. Повышение частоты тестов приводило к формированию обучаемости, от чего сигналы от стимулов становились более устойчивыми и быстрыми. Это говорит о том, что таламические синапсы в неокортексе кодируют предыдущий опыт подопытного.
Однако существует разница между стимулами, получаемыми мозгом. Некоторые стимулы вызывают определенную реакцию, независимо от опыта. К примеру, чем громче звук, тем легче он привлекает внимание человека или подопытной мыши. Однако это функция низкого уровня, имеющая мало общего с вышеописанным опытом. Тем не менее, как оказалось, существует ранее неизвестный механизм, который может точно кодировать информацию, идентифицируя специализированный тип нейрона в самом верхнем слое неокортекса как динамический вход этих нисходящих сигналов.
Следовательно, таламический вход в сенсорный неокортекс является ключевым источником информации о прошлых событиях, связанных с сенсорными стимулами.
Важность данного открытия заключается в том, что в ходе развития определенных заболеваний мозга (аутизм, шизофрения и т.д.) кодирование сенсорной информации, связанной с накопленным опытом, нарушается. Зная, какие именно механизмы задействованы в этом процессе, а также как именно они протекают, ученые могут разработать методику восстановления нарушений, вызванных болезнью.
Важность мозга для работы нашего организма нельзя отрицать, однако на данный момент, несмотря на технологический прогресс, нам по-прежнему известно о нем далеко не все. Чем больше будет проводиться подобного рода исследований, тем лучше мы будем понимать наш собственный организм.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята. 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Похожие статьи
 Что почитать на выходных: «Теория каст и ролей» Алекса Крола
Что почитать на выходных: «Теория каст и ролей» Алекса Крола Эволюция профессий: что важно знать современным айтишникам о прошлом и будущем труда
Эволюция профессий: что важно знать современным айтишникам о прошлом и будущем труда Эксклюзивное интервью Хабра с Ником Бостромом
Эксклюзивное интервью Хабра с Ником Бостромом Три секретами европейских валют
Три секретами европейских валют Видеосвязь больше не гарантия: как дипфейки подрывают доверие в бизнесе
Видеосвязь больше не гарантия: как дипфейки подрывают доверие в бизнесе Как мыслит языковая модель
Как мыслит языковая модель Обычные телескопы способны обнаружить межзвездные лазерные маяки внеземных цивилизаций
Обычные телескопы способны обнаружить межзвездные лазерные маяки внеземных цивилизаций Здравый смысл против релятивизма
Здравый смысл против релятивизма