
Современные роботы отличаются от своих предшественников не только улучшенными интеллектуальными способностями, но и маневренностью. Стоит вспомнить видео Boston Dynamics, где инженеры пинают и толкают несчастного робота, демонстрируя его умение сохранять стабильность или восстанавливаться после падения. Но во всех этих роликах робот цел, то есть, как бы мрачно это не звучало, все его конечности на месте. В дикой природе существуют механизмы восстановления, помогающие различным видам сохранять стабильность движения в случае серьезной травмы. Ученые из университета штата Пенсильвания (США) решили изучить этот вопрос на примере фруктовой мошки, способной летать даже при потере 40% своих крыльев. Как именно мошки сохраняют мобильность после таких травм, какова динамика их движения до и после травмы, и как полученные данные можно применить в робототехнике? Ответы на эти вопросы мы найдем в докладе ученых.
Основа исследования
Одним из основных аспектов любой биологической системы, будь то животное или растение, является ее умение компенсировать возмущения внутреннего или внешнего происхождения. Возмущения окружающей среды могут иметь серьезные последствия для выживания, в некоторых случаях вызывая травмы, которые ограничивают подвижность. К примеру, у членистоногих повреждения могут возникать в результате дефектов линьки, болезней, хищничества, агрессивного поведения представителей своего же вида и т. д. Исследование природных популяций членистоногих показало, что у 40% определенных видов членистоногих отсутствует по крайней мере один придаток целиком. Следовательно, возмущения могут быть скорее нормой, чем исключением.
Если говорить о летающих насекомых, то для них важнейшим атрибутом являются крылья. С их помощью насекомые могут избежать атаки хищника, быстрее найти партнера или пищу, расширить ареал своего обитания и т. д. Повреждение крыльев напрямую влияет на смертность таких существ, потому они обладают рядом адаптационных механизмов, способных компенсировать травмированные крылья. Насекомые, в отличие от птиц и летучих мышей, не могут восстанавливать повреждения крыльев, потому они столь сильно нуждаются в стратегиях компенсаторного контроля со стороны сопряженных нейронных и механических систем.
Одним из ярких примеров устойчивости к естественным повреждениям крыльев являются мухи. С инженерной точки зрения мухи могут реализовывать контроль над своим полетом при наличии повреждений двумя разными методами: с и без внутренних изменений нервной системы. Оба метода могут работать в сопряжении, но то, как каждый из них реализуется, играет большое значение для целостного понимая концепции адаптации.
В рассматриваемом нами сегодня труде ученые применили теоретическую основу контроля, чтобы показать, как сенсорная обратная связь позволяет мухам компенсировать повреждение крыльев в полете. Чтобы количественно оценить, как повреждение крыла влияет на летные характеристики, ученые количественно оценили моторику мух с неповрежденными и односторонне поврежденными крыльями.
Результаты исследования
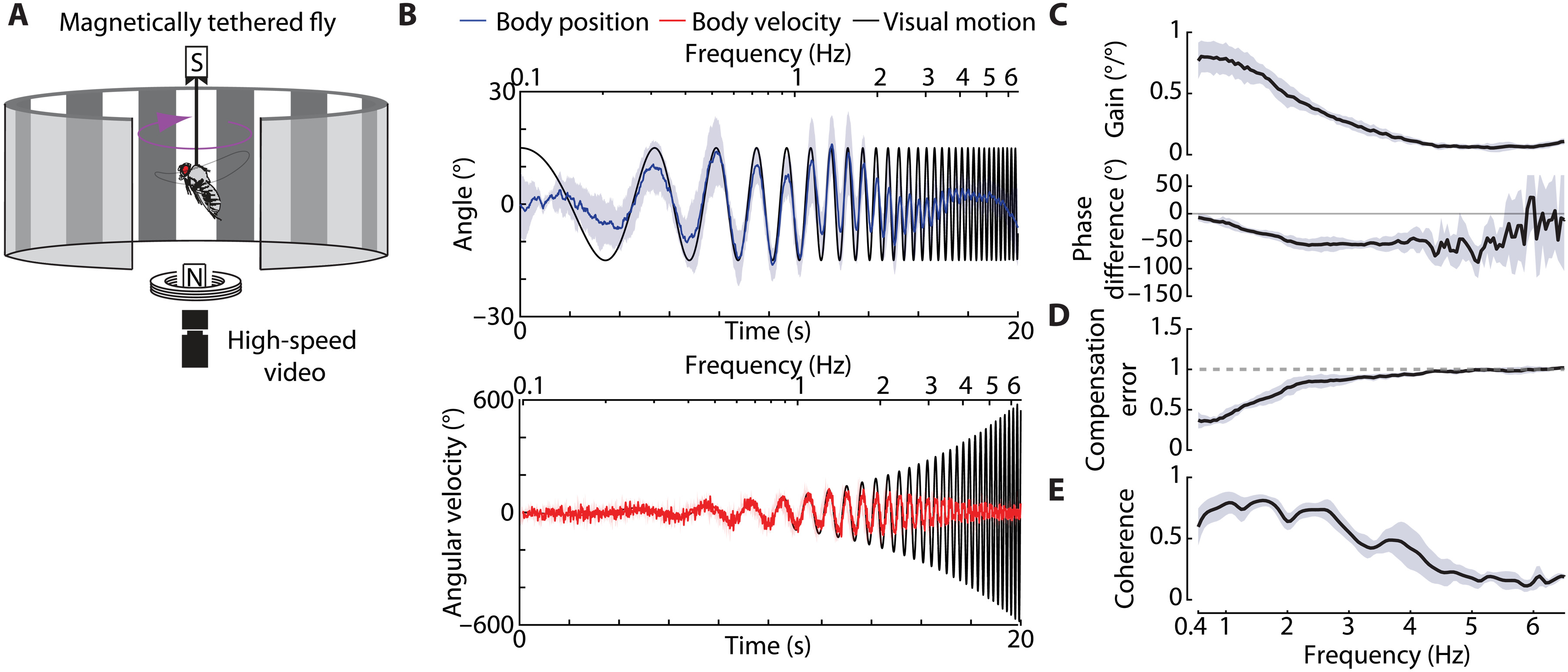
Чтобы изучить влияние повреждения крыла на летные характеристики, ученые сначала попытались определить свойства оптомоторного рефлекса рыскания тела у неповрежденных мух посредством визуальных стимулов, направленных на особей, магнитно закрепленных в камере наблюдения (1А).
Изображение №1
Визуальный стимул представлял собой чередующиеся полосы разного цвета (наподобие штрих-кода), на вращающейся вокруг мухи ленте. Частота смены полос увеличивалась логарифмически с 0.1 до 6.5 Гц в течение 20 секунд и демонстрировалась с постоянной амплитудой 15°, представляя пиковые скорости от 9° до 612° в секунду. Данный диапазон скоростей находятся в пределах визуальной полосы пропускания оптомоторного ответа дрозофилы (1B). Проще говоря, мухи были способны визуально фиксировать изменения стимула и реагировать на него.
Этот стимул вызывал оптомоторный ответ на всех частотах, но ответ был прежде всего настроен на низкие частоты (пиковые скорости от 38° до 188° в секунду), представляющие область с самым высоким усилением и когерентностью, а также наименьшую ошибку компенсации (1C—1E). На низких частотах коэффициент усиления с обратной связью был близок к единице, а разность фаз была приблизительно равна нулю, что свидетельствует о высокой эффективности компенсации. Однако по мере увеличения частоты стимула как усиление, так и разность фаз быстро уменьшались, что соответствовало снижению эффективности компенсации.
Выше 4 Гц (пиковая угловая скорость 377 °/с) усиление и когерентность были близки к нулю, а разность фаз становилась чрезвычайно зашумленной. Это позволяет предположить, что мухи почти не реагируют на визуальное движение в этом более высокочастотном диапазоне.
Ниже 0.4 Гц (пиковая угловая скорость 28 °/с) мухи реагировали на каждый шаг отдельных пикселей светодиодной арены. Таким образом, стимул не вызывал плавного движения и в целом имел низкую когерентность, что еще больше указывало на нелинейность (1C). Потому дальнейшие опыты проводились на частотах выше этого порога.

Изображение №2
Далее было проведено изучение того, как одностороннее повреждение крыла может повлиять на работу оптомоторной реакции (хордовый срез левого крыла с диапазоном потери площади от 10 до 40%; 2А).
Видео №1: наблюдение за мухой с неповрежденными крыльями.
В ходе наблюдений за травмированными мухами было отмечено, что они медленно дрейфовали в направлении поврежденного крыла (2B и 2C).
Видео №2: наблюдение за мухой с поврежденным крылом.
Особи без повреждений также демонстрировали дрейф, но он был слабым и случайным в аспекте направления (2C).
Чтобы разделить наблюдаемый дрейф и летные характеристики после повреждения крыла, был проведен анализ угловой скорость обеих групп. Обе группы продемонстрировали одинаковые результаты в диапазоне частот стимула. Однако усиление с обратной связью существенно упало на частоте 3.7 Гц для обеих групп мух, что свидетельствует о фильтрации нижних частот (2D). Разность фаз постепенно увеличивалась одинаковым образом для обеих групп с увеличением частоты, снова указывая на фильтр нижних частот. Статистическое сравнение производительности неповрежденных и поврежденных мух показало значительную разницу в усилении на всех частотах, но не в разнице фаз (2D). Статистическое сравнение производительности неповрежденных и поврежденных мух показало значительную разницу в усилении на всех частотах, но не в разнице фаз. Все это указывает на незначительное снижение эффективности стабилизации полета у мух с поврежденным крылом по сравнению с мухами с целыми крыльями.
Чтобы измерить, как степень повреждения крыльев повлияла на летные характеристики, мух разделили на категории по объему повреждений крыльев. Всего было три группы: 30%, 20-30% и менее 20%. Статистический анализ не выявил существенной разницы между усилением и разностью фаз трех групп для всех частот. Эти результаты показывают, что мухи, потерявшие около 40% общей площади крыльев, вели себя так же, как и мухи, потерявшие только 10%.
Далее было проведено дополнительное наблюдение за мухами с и без повреждений в условиях тестовой камеры. В ходе наблюдений ученые определяли стабильных и нестабильных особей. Под нестабильными подразумевались те, что непрерывно вращались и тряслись, а потому не могли стабилизировать статическое положение. Из 78 мух с поврежденным крылом 36 мух были нестабильны (46%), а из 50 мух с целыми крыльями 20 были нестабильны (40%).
Статистический анализ показал, что эти две пропорции не были статистически значимыми. Следовательно, мухи могут легко компенсировать повреждения даже в зафиксированном состоянии, а доля стабильных мух не отличалась между неповрежденными и поврежденными состояниями.
Итоговый вывод заключается в том, что особи, потерявшие 40% или менее от площади крыльев, не демонстрировали потери в контроле положения тела во время полета.
Хотя и было обнаружено общее снижение эффективности вращения тела после повреждения крыла, мухи все же могли компенсировать снижение вращения тела, регулируя движения головы. Движения головы у дрозофилы имеют меньшую зрительно-моторную задержку, чем движения крыльев, и могут уменьшить скольжение сетчатки примерно на 60%. Таким образом, голова может легко компенсировать асимметрию в создании крутящего момента крыла.
Отслеживая ориентацию головы зафиксированных мух относительно фиксированной вертикальной оси, ученые измерили движение головы у мух с целыми и поврежденными крыльями. Анализ показал, что движение головы мух с поврежденным крылом не увеличивалось из-за уменьшения движения тела.
Чтобы определить, распространяется ли снижение моторики на другие классы стимулов, біла измерена реакция тела мухи на панораму, движущуюся с постоянной скоростью. Подтверждая предыдущий опыт, мухи с целыми крыльями компенсировали движущийся фон, почти совпадая с его скоростью, давая усиление, близкое к единице, где усиление определяется как отношение скорости мухи к скорости фона (2E). Как и в предыдущем опыте, каких-либо значимых отклонений в динамике движений мух с поврежденным крылом обнаружено не было.
Одностороннее повреждение крыла незначительно снижает эффективность стабилизации, и мухи могут активно компенсировать асимметричный крутящий момент, изменяя кинематику своего крыла.
К примеру, в свободном полете Drosophila hydei, более крупный вид плодовой мухи, чем Drosophila melanogaster, изменяет кинематику крыла после одностороннего повреждения крыла. D. Hydei увеличивает частоту взмахов крыла и асимметрично регулирует движение крыла, чтобы противодействовать дисбалансу крутящего момента по крену из-за односторонней потери аэродинамической поверхности. Однако степень, в которой эти компенсационные стратегии управляются активными или пассивными механизмами, остается неясной.
Например, асимметрия между углами хода левого и правого крыльев может возникать из-за разности сил сопротивления из-за односторонней потери аэродинамической поверхности крыла. Кроме того, уменьшение инерции крыла может сопровождаться сопутствующим увеличением частоты взмахов из-за увеличения резонансной частоты механической системы крыло-грудь и, следовательно, может не требовать активного контроля со стороны нервной системы. Исследования этого вопроса показали, что частота колебаний асинхронной мышцы близка к резонансу без какой-либо обратной связи со стороны нервной системы.

Изображение №3
Авторы рассматриваемого нами труда решили определить потенциальное влияние пассивных аэродинамических и механических воздействий на компенсационные стратегии полета. Для этого было проведено сравнение WBA (двумерная проекция угла взмаха) и частоту взмахов мух при жесткой и магнитной фиксации в тестовой камере. Ученые объясняют, поскольку эти две парадигмы (разные типы фиксации особей) обеспечивают разные топологии управления (открытый или замкнутый контур), сравнение поведения в полете между этими парадигмами может дать уникальное представление о вкладе сенсорной обратной связи в управление полетом (3А).
Ученые пришли к выводу, что если относительные изменения WBA между интактными и поврежденными мухами были одинаковыми при наличии (магнитная привязка) и дефиците (жесткая привязка) зрительной референции и механо-сенсорной обратной связи тела, то увеличение WBA лучше всего объяснялось бы пассивной механикой в отличие от активной компенсации. Точно так же, если бы частота взмахов крыльев была одинаковой в обеих парадигмах, то пассивная механика могла бы быть лучшим объяснением изменений в частоте взмахов крыльев.
Чтобы выявить вклад активных и пассивных механизмов, сначала было проведено сравнение WBA в жесткой и магнитной фиксации, когда мухам предъявляли статический визуальный рисунок, тем самым моделируя условия зависания. Также была выполнена количественная оценка изменения WBA (ΔWBA) между левым и правым крылом. Как в жесткой, так и в магнитной фиксации одностороннее повреждение крыла значительно увеличивало ΔWBA между целыми и поврежденными мухами (3B). Однако ΔWBA при магнитной фиксации было значительно больше, чем при жесткой. Вместе эти результаты показывают, что пассивная аэродинамика и визуальная обратная связь способствовали увеличению ΔWBA.
Чтобы показать, как визуальная обратная связь модулирует ΔWBA, были количественно определены левая и правая WBA по отдельности как в жесткой, так и в магнитной фиксации (3C и 3D). WBA у мух с поврежденным крылом различался между парадигмами. Это позволяет предположить, что модуляции WBA поврежденного крыла управляются как пассивными, так и активными механизмами.
Вместе эти результаты показывают, что как пассивная механика, так и сенсорная обратная связь лежат в основе контроля полета после одностороннего повреждения крыльев.
У мух на жесткой фиксации частота взмахов крыльев зависела от степени повреждения (3E). Удаление части крыла приводило к уменьшению инерции колебательной системы, состоящей из двух крыльев и торакса (груди). Ранее сообщалось, что частота взмахов крыльев зависит от инерции крыла, как и предсказывает линейная модель пружинно-массового демпфера.
Данная модель могла объяснить лишь незначительную дисперсию, но вот высокая дисперсия частоты взмахов крыльев связана с тем, что она находится под активным контролем и с тем, что мухам не нужно взмахивать крыльями в резонансе с грудной клеткой. Мухи на жестком креплении могли активно модулировать частоту взмахов крыльев из-за изменений в зрительной и гироскопической обратной связи.
Видео №3: сравнение динамики целого и поврежденного крыла.
Другим объяснением данной картины может быть тот факт, что различия в кинематике полета отражают различные стратегии управления полетом отдельных мух. Следовательно, изменения частоты взмахов после повреждения крыла, вероятно, являются проявлением как пассивных, так и активных механизмов.
Точно так же амплитуда взмахов поврежденного (левого) крыла увеличивалась с увеличением степени повреждения, хотя наблюдалась значительная вариабельность (3F).
Объединив данные о полете с результатами математического моделирования, можно предположить, что как активный визуальный контроль, так и пассивная механика лежат в основе стратегий полета для компенсации повреждений крыла.

Изображение №4
Во время наблюдений также было установлено, что мухи с поврежденными крыльями смещали положение своего брюшка примерно на 9° в сторону неповрежденного крыла, что указывает на активную компенсацию повреждений (4А).
Чтобы определить, может ли абдоминальная реакция (смещение брюшка) быть вызвана невизуальными ощущениями, например, механосенсорной обратной связью от поврежденного крыла, ученые повторили опыты с жесткой/магнитной фиксацией.
Смещение брюшка у интактных (без повреждения крыла) и поврежденных мух как в магнитной, так и в жесткой фиксации значительно отличалось (4A).
Абдоминальная реакция поврежденных мух в жесткой фиксации масштабировалась с повреждением крыла, которое само масштабировалось с ΔWBA. Это может означать, что повреждение крыла вызывает компенсаторную брюшную реакцию невизуального происхождения (4B).
Вместе эти результаты подтверждают гипотезу о том, что изменение положения брюшка управляется механосенсорной рефлекторной обратной связью между крыльями и брюшком.
В ходе исследования ученые решили проверить еще одну гипотезу, которая указывает на адаптивный контроль, который позволяет системе изменять коэффициенты усиления контроллера для достижения аналогичной производительности с обратной связью, например, после изменений в теле (объекте), которые могут быть вызваны повреждением. Отличительной чертой адаптивных систем управления является то, что параметры контроллера не являются фиксированными: они изменяются в ответ на изменения динамики системы и неопределенностей.
Изображение №5
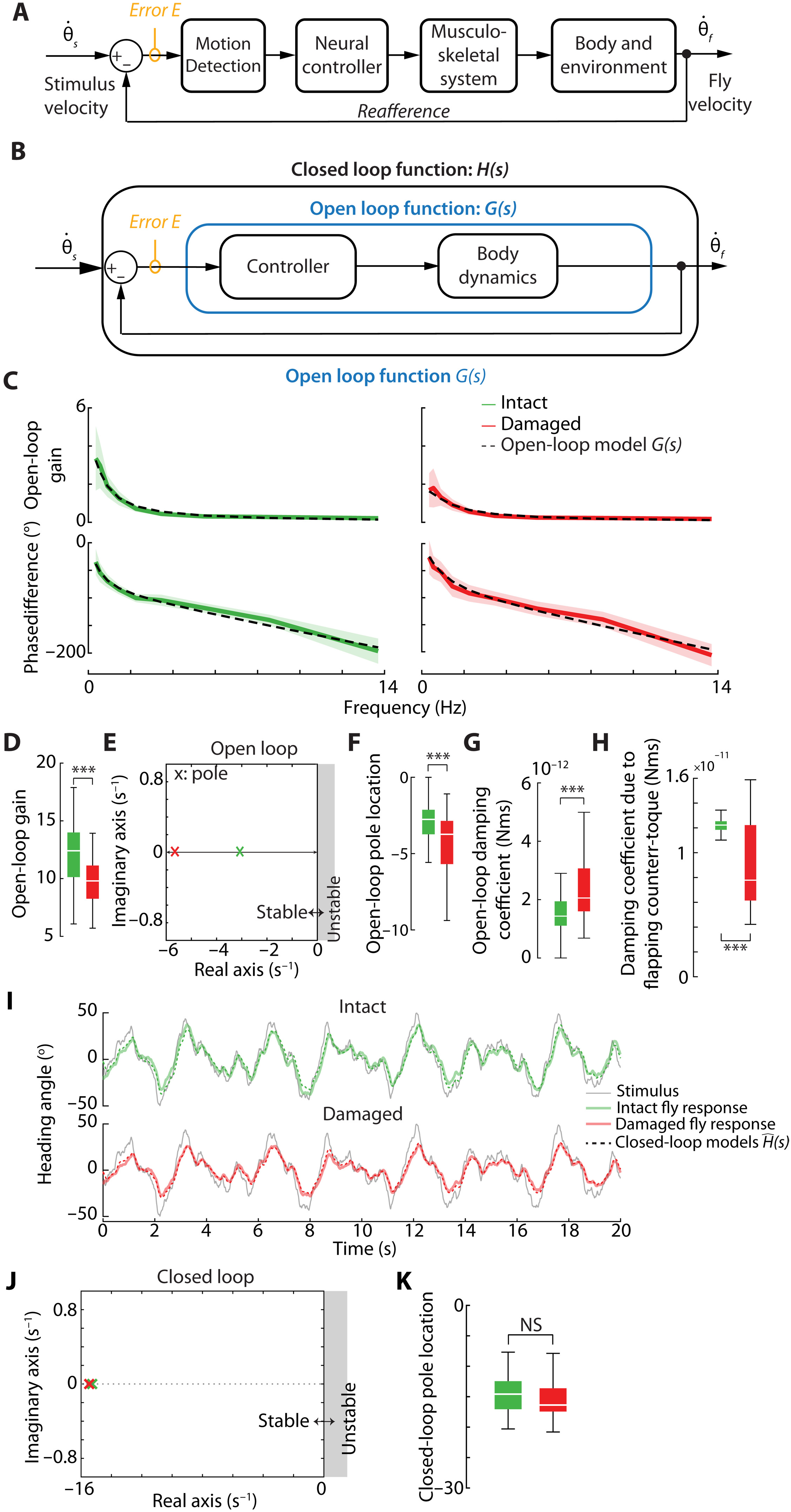
Чтобы определить, может ли нейронный контроллер мух быть адаптивным к различным внешним возмущениям системы (т. е. параметры контроллера изменяются из-за повреждения крыла), ученые вывели функции параметрической частотной характеристики (FRF от frequency response function) отдельных мух. Используя полученные FRF, они оценили параметры внутреннего нейронного контроллера и динамики тела (5А). В итоге была проведена оценка передаточной функции без обратной связи (G).
Оценивая числитель и знаменатель G, можно получить критическое представление о том, как мухи компенсируют повреждение крыла. Числитель объединяет усиление нейронного контроллера и статическое усиление тела, называемое усилением без обратной связи. Знаменатель определяет положение полюса и, тем самым, устойчивость целых и поврежденных мух (5D). Карта полюс-ноль также выявила разные параметры между обеими системами без обратной связи, несмотря на то, что обе системы были стабильными (5E). Поврежденные мухи в целом имели более высокий коэффициент демпфирования по сравнению с неповрежденными мухами (5F), что могло быть связано с увеличением амплитуды и частоты взмахов крыльев.
Вместе эти результаты показывают, что повреждение крыла не только изменяет характеристики стабилизации рыскания у мух, но также изменяет связанную динамику тела и нейронный контроллер, используемый для стабилизации взгляда.
Дополнительный сравнительный анализ показал, что снижение эффективности полета у поврежденных мух имеет преимущество в поддержании стабильности на уровне, подобном интактным мухам. Другими словами, достигается компромисс, за счет которого мухи способны лететь дальше, несмотря на повреждения.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня исследовании ученые уделили внимание фруктовым мошкам и тому, как они продолжают летать, несмотря на весьма существенные повреждения крыла. Эти существа способны летать, даже если у них отсутствует порядка 40% от общей площади крыльев. Следовательно, возникает вопрос — как они компенсируют это повреждение.
В ходе наблюдений было установлено, что мухи компенсировали травму за счет увеличения частоты взмахов поврежденного крыла и уменьшения частоты целого. Этот на первый взгляд простой процесс достигается за счет модуляции нервной системы, что позволяет мухам точно настраивать свой полет даже при наличии повреждений.
Ученые отмечают, что при повреждении крыла обычно страдает как производительность, так и устойчивость. Мухи в случае повреждения крыла готовы снизить производительность полета, дабы поддерживать стабильность на желаемом уровне.
Данное исследование предполагает, что плодовые мошки, имеющие порядка 200 тысяч нейронов, используют сложную и гибкую систему управления моторикой, позволяющую им адаптироваться и выживать после травмы. Ученые отмечают, что найденная в ходе система контроля у мух превосходит таковые по сложности даже у самых современных летающих роботов. Вполне очевидно, что для инженеров-робототехников подобные открытия крайне полезны. Учитывая, что летающие насекомые всегда были одним из источников вдохновения для летающих роботов, понимание того, как именно эти биологические системы справляются с нестандартными ситуациями, может помочь усовершенствовать роботов, сделав их более маневренными, долговечными и полезными.
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


