
Клин клином: использование вируса PIV5 для создания вакцины против MERS-CoV и, возможно, против SARS-CoV-2
Учитывая сложившиеся обстоятельства, вовсе неудивительно, что научное сообщество сконцентрировало все свои силы на поиск и разработку методик и инструментов для борьбы с распространяющимся вирусом. Сегодня мы рассмотрим исследование, в котором ученые из Американского общества по микробиологии (ASM) описывают вполне рабочую вакцину против вируса MERS, начавшего свое распространение в 2012 году. По мнению ученых, их разработка может помочь в создании вакцины против ныне бушующего SARS-CoV-2. В чем особенность созданной вакцины, из чего она была создана, и насколько эффективно она выполняет свою задачу? Об этом мы узнаем из доклада ученых. Поехали.
Основа исследования
MERS или ближневосточный респираторный синдром — воспалительное заболевание дыхательных органов, который развивается ввиду заражения вирусом рода Betacoronavirus из знакомого нам сейчас подсемейства Coronavirinae.
Снимок MERS-CoV (желтый цвет).
Источником вируса MERS-CoV опять же являются летучие мыши. Первые случаи заражения среди людей были зафиксированы еще в 2012 году на территории Аравийского полуострова, однако уже к 2015 году вирус добрался и до стран Восточной Азии и Европы. По последним данным число подтвержденных случаев инфицирования MERS-CoV составляет 2494 человека, среди которых умерло 858 (данные от 04.02.2020). Данные разнятся от источника к источнику, однако отчетливо видно, что степень распространения у MERS-CoV не так велика, как у SARS-CoV-2. Тем не менее ближневосточный вирус также обладает достаточно высокой смертностью — около 35%. Именно этот показатель беспокоит больше всего.
MERS-CoV представляет собой РНК-вирус с положительной цепью*, у которого проникновение в клетки-мишени осуществляется за счет S-белка вирусной оболочки*.
РНК-вирус с положительной цепью* — положительная вирусная РНК. Это подразумевает, что конкретная последовательность вирусной РНК может быть непосредственно транслирована в вирусные белки (например, белки, необходимы для репликации вируса). Следовательно, в РНК-вирусах с положительной цепью вирусный РНК-геном можно считать вирусной мРНК (матричная РНК) и он может быть немедленно транслирован клеткой-хозяином.
Вирусная оболочка* — дополнительная оболочка, покрывающая внешнюю оболочку (капсид) многих вирусов.
S-белок состоит из субъединицы S1, ответственной за связывание с рецептором вируса, дипептидилпептидазой 4 (DPP4 или CD26) через рецептор-связывающий домен (RBD от receptor-binding domain), а также из субъединицы S2, которая обеспечивает слияние мембран.
Исследователи напоминают, что на данный момент вакцины против MERS-CoV нет, есть только ряд разработок, эффективность которых достаточно сомнительна. Ученые считают, что их труд изменит эту ситуацию.
В данном исследовании ученые решили обратить свое внимание на еще один вирус — PIV5. PIV или вирусы парагриппа (ПГ) насчитывают пять разновидностей, три из которых вызывают заболевания у человека (от ПГ-1 до ПГ-3) и два, которые для человека не опасны (ПГ-4 и ПГ-5).
ПЭМ снимок вируса парагриппа.
ПГ-5 является членом рода Rubulavirus из семейства Paramyxoviridae, в состав которого входят вирус эпидемического паротита (MuV или «свинка») и вирус парагриппа человека типа 2 (ПГ-2) и типа 4 (ПГ-4). ПГ-5 кодирует восемь известных вирусных белков. Нуклеокапсидный белок (NP), фосфопротеин (P) и белок РНК-полимеразы (L) важны для транскрипции* и репликации* генома вирусной РНК.
Транскрипция* — процесс синтеза РНК с применением ДНК в качестве матрицы; перенос генетической информации с ДНК на РНК.
Репликация* — процесс создания двух дочерних молекул ДНК на основе родительской ДНК.
Ученые считают, что ПГ-5 является отличным кандидатом в вирусные векторы* для будущей вакцины. ПГ-5 вполне безопасен, несмотря на свою высокую степень заразность, что может сыграть на руку при создании эффективной вакцины.
Вирусные векторы* — инструменты для доставки генетического материала в клетки-мишени.
Кроме того, ПГ-5 не имеет ДНК фазы в своем жизненном цикле, что позволяет избежать возможных непреднамеренных генетических модификаций ДНК клетки-хозяина путем рекомбинации или вставки. Также структура ПГ-5 достаточно стабильная, в отличие от РНК-вирусов с положительной цепью. Подтверждением тому является созданный рекомбинантный ПГ-5, экспрессирующий ген F, для респираторно-синцитиального вируса, когда ген F поддерживался в течение более 10 поколений.
Еще одним преимуществом ПГ-5 является его экономичность, так как данный вирус можно выращивать вплоть до 8х108 БОЕ/мл (БОЕ — бляшкообразующих единиц).
Ранее ПГ-5 уже использовался в качестве вирусного вектора при работе с вакцинами против респираторно-синцитиального вируса и бешенства. Во время опытов на мышах было установлено, что вектор ПГ-5, экспрессирующий NA (нейраминидаза, входящая в состав оболочки) вируса гриппа, приводит к полной иммунизации подопытных, т.е. спустя 4 дня после заражения не было ни одного случая смерти или даже заражения. А вот ПГ-5, экспрессирующий NP (нуклеопротеин), защищает 100% подопытных мышей от смертельного для них штамма вируса гриппа H1N1.
Основываясь на столь позитивных результатах ранее проведенных исследований, ученые решили использовать ПГ-5 в качестве вирусного вектора, экспрессирующего S-белок вируса MERS. Как и ранее, все опыты проводились на мышах.
Результаты исследования
Исследователи заранее провели внедрение HA (гемагглютинин) гена вируса гриппа A в разные места в геноме ПГ-5 и обнаружили, что вставки в SH (малый гидрофобный) и HN (гемагглютинин-нейраминидаза) дают лучший иммунный ответ.
Гемагглютинин и нейраминидаза вируса гриппа* — поверхностные антигены вируса, обеспечивающие возможность присоединяться к клеткам-мишеням.
Учитывая это, было решено внедрить полноразмерный S ген вируса MERS на пересечении SH и HN. Плазмиду*, содержащую полноразмерную кДНК* ПГ-5 с внедренным S геном в месте соединения SH и HN, конструировали с использованием стандартных методов молекулярного клонирования (1А).
Изображение №1
Плазмида* — небольшая молекула ДНК, отделенная от хромосом и способная к самостоятельной репликации.
кДНК* — комплементарная ДНК, т.е. ДНК, синтезированная на матрице зрелой мРНК (матричная РНК) в реакции, катализируемой обратной транскриптазой.
Плазмиду трансфицировали* в BHK* клетки вместе с плазмидами, экспрессирующими РНК полимеразу T7, NP, P и L вируса ПГ-5 (PIV5).
Трансфекция* — внедрение нуклеиновой кислоты в клетки невирусным методом.
BHK клетки* (baby hamster kidney) — клетки соединительной ткани (фибробласт) почек хомяка, чувствительные ко многим вирусам, воздействующим на человека. BHK клетки используются для трансформации, а также для стабильных и временных трансфекций.
В результате был получен инфекционный вирус PIV5-MERS-S, который далее прошел очистку и был размножен в большом количестве в MDBK клетках* для дальнейшего анализа.
MDBK клетки* (Madin-Darby canine kidney cells) — клетки почки собаки, впервые выделенные в 1958 году учеными S. H. Madin и N. B. Darby. Данные клетки используются в исследованиях в качестве модельных клеток млекопитающих.
Геном вируса секвенировали и подтвердили, что он содержит желаемую последовательность входной ДНК. Далее необходимо было проверить экспрессию S белка в клетках, инфицированных PIV5-MERS-S. Для этого клетки инфицировали при разных значениях MOI*, после чего они были лизированы (растворены) для иммуноблоттинга* с использованием анти-S антитела.
MOI* (multiplicity of infection) — кратность заражения, т.е. отношение агентов (вируса) к мишеням для инфицирования (клеткам).
Иммуноблоттинг* — высокочувствительный метод выявления белков, основанный на сочетании электрофореза и иммуноферментного или радиоиммунного анализа.
Анализ показал наличие как полного S белка, так и S2 фрагментов, что указывает на успешность его внедрения (1В). Экспрессия S белка в PIV5-MERS-S-инфицированных клетках была дополнительно подтверждена иммунофлуоресцентным анализом (1С).
Ученые отмечают, что PIV5-MERS-S вызывал массивное образование синцития* в клетках Vero*. Помимо этого, кинетика роста PIV5-MERS-S и обычного ПГ-5 (PIV5) была очень похожа (1D).
Синцитий* — тип ткани, в которой клетки не полностью разделены, т.е. имеются участки цитоплазмы с ядрами, которые связаны между собой плазмодесмами (цитоплазматическими мостиками).
Vero* — используемая для культивирования линия клеток, полученная из эпителия почки африканской зеленой мартышки.
Изображение №2
Иммунизация PIV5-MERS-S генерирует нейтрализующие антитела и опосредованный Т-клетками иммунитет. Чтобы определить, может ли PIV5-MERS-S генерировать иммунные ответы, мышей иммунизировали однократной дозой PIV5-MERS-S или контрольным вирусом PIV5-GFP при 104 БОЕ или 106 БОЕ на особь через интраназальный путь.
В то время как обе дозы вызывали ответы антител, нейтрализующие титры были скромными при 1:64 и 1:128 для 104 и 106 доз соответственно (2А и 2В).
Ученые напоминают, что мыши видов C57BL/6 и BALB/c после иммунизации генерируют Th1- и Th2-доминантные иммунные ответы, соответственно.
Th (Т-хелперы) — T-лимфоциты, усиливающие адаптивный иммунный ответ (приобретенный иммунитет).
Th1 — содействуют развитию клеточного иммунного ответа, активируя макрофаги.
Th2 — активируют В-лимфоциты, что помогает развитию гуморального иммунного ответа.
Одна доза PIV5-MERS-S (106 БОЕ) приводила к титру нейтрализующих антител вплоть до 1:2000 у мышей BALB/c (2C), что согласуется с Th2-доминантным ответом у мышей данного вида.
Для оценки первичного ответа CD8* Т-клеток, генерируемого иммунизацией PIV5-MERS-S, мышей hDPP4-KI интраназально иммунизировали PIV5-MERS-S (104 БОЕ).
CD8* (кластер дифференцировки 8) — трансмембранный гликопротеин, работающий как корецептор Т-клеточных рецепторов.
Спустя четыре недели был произведен забор для исследования наличия/отсутствия MERS-CoV-специфических легочных резидентных CD8 Т-клеток (3А).
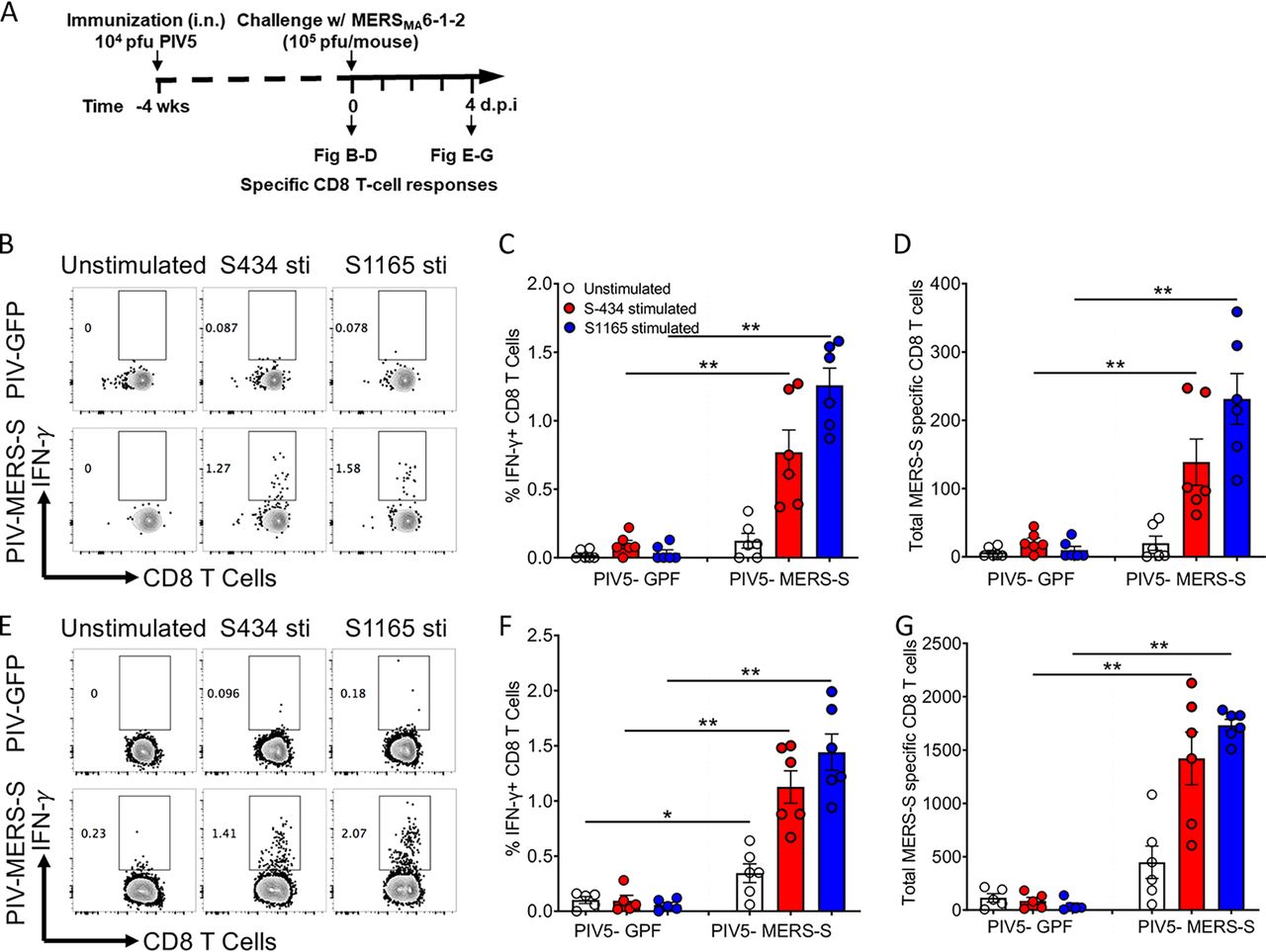
Изображение №3
Как показано на 3B—3D, наблюдалось значительное увеличение процента и общего количества CD8-IFN-клеток в легких у мышей, иммунизированных PIV5-MERS, по сравнению с инфицированными вирусом PIV5-GFP.
Для оценки ответной реакции MERS-S-специфичных CD8 Т-клеток было проведено дополнительное внедрение MERS-CoVMA(104 БОЕ). В результате было видно значительное увеличение ответа CD8 Т-клеток уже на четвертый день по сравнению с мышами, инфицированными контрольным PIV5-GFP (3E—3G).
Также наблюдалось 10-кратное увеличение MERS-S-специфических CD8 T-клеток по сравнению с первичным ответом CD8 T-клеток (3B—3G).
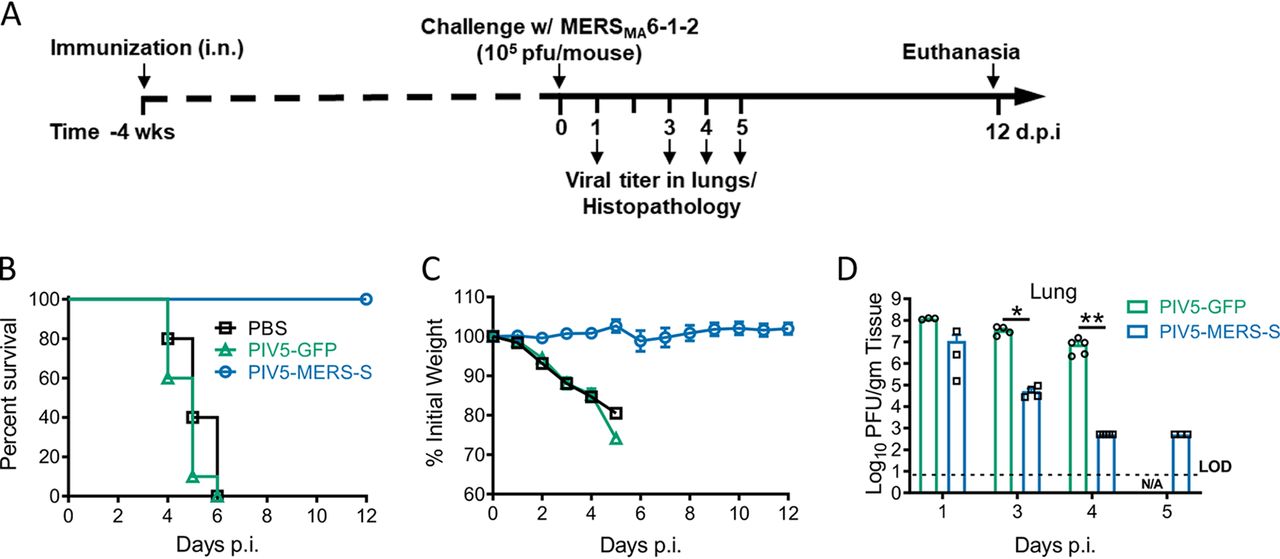
Изображение №4
В результате PIV5-MERS-S иммунизация предотвращает летальный исход инфицирования у мышей. Чтобы определить эффективность PIV5-MERS-S в профилактике или модификации вируса MERS-CoV, мышей hDPP4 KI иммунизировали PIV5-MERS-S (104 БОЕ) интраназальным путем.
Через 4 недели после иммунизации мышей заражали MERS-S, модифицированным под более активное воздействие на мышей (4А). Все мыши, иммунизированные PIV5-MERS-S, пережили это смертельное заражение, при этом наблюдалась незначительная потеря веса (4B и 4C). В противовес этому, все мыши, инфицированные контрольным PIV5-GFP или PBS, умерли после заражения. Это указывает на то, что PIV5-MERS-S полностью защищал мышей от летального заражения.
Несмотря на то, что PIV5-MERS-S иммунизированные мыши имели более высокую скорость очищения легких от вируса, данный метод не обеспечивал полного стерилизующего иммунитета (4D).
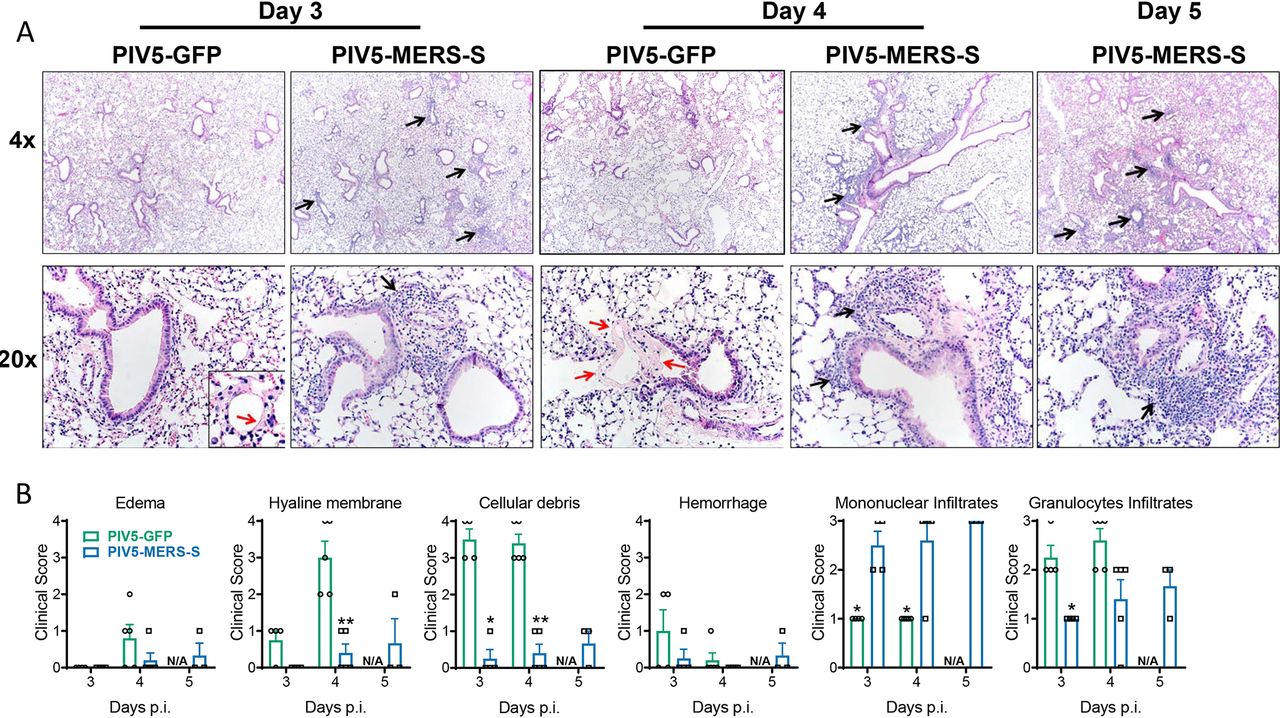
Изображение №5
Гистопатологические исследования легких после заражения показали, что у мышей, иммунизированных PIV5-MERS-S, было значительно меньше клеточного дебриса* и больше мононуклеарных инфильтратов (5А и 5В).
Клеточный дебрис* – остаток клетки, окруженный плазматической мембраной, который фагоцитируется макрофагами.
Также была выявлена сильная клеточная инфильтрация лейкоцитов (в основном мононуклеарных клеток) и меньше повреждений (отек, гиалиновые мембраны, некротический клеточный дебрис и т.д.).
На заключительном этапе исследования ученые провели оценку защитных ответов, вызванных PIV5-MERS-S или инактивированным MERS-CoV. Для этого одних мышей иммунизировали PIV5-MERS-S (104 БОЕ) или PBS, тогда как других иммунизировали УФ-инактивированным MERS-CoV.
В то время как PIV5-MERS-S обеспечивал 100% защиту от летального заражения, инактивированный MERS-CoV защищал лишь 25% мышей (изображение ниже).
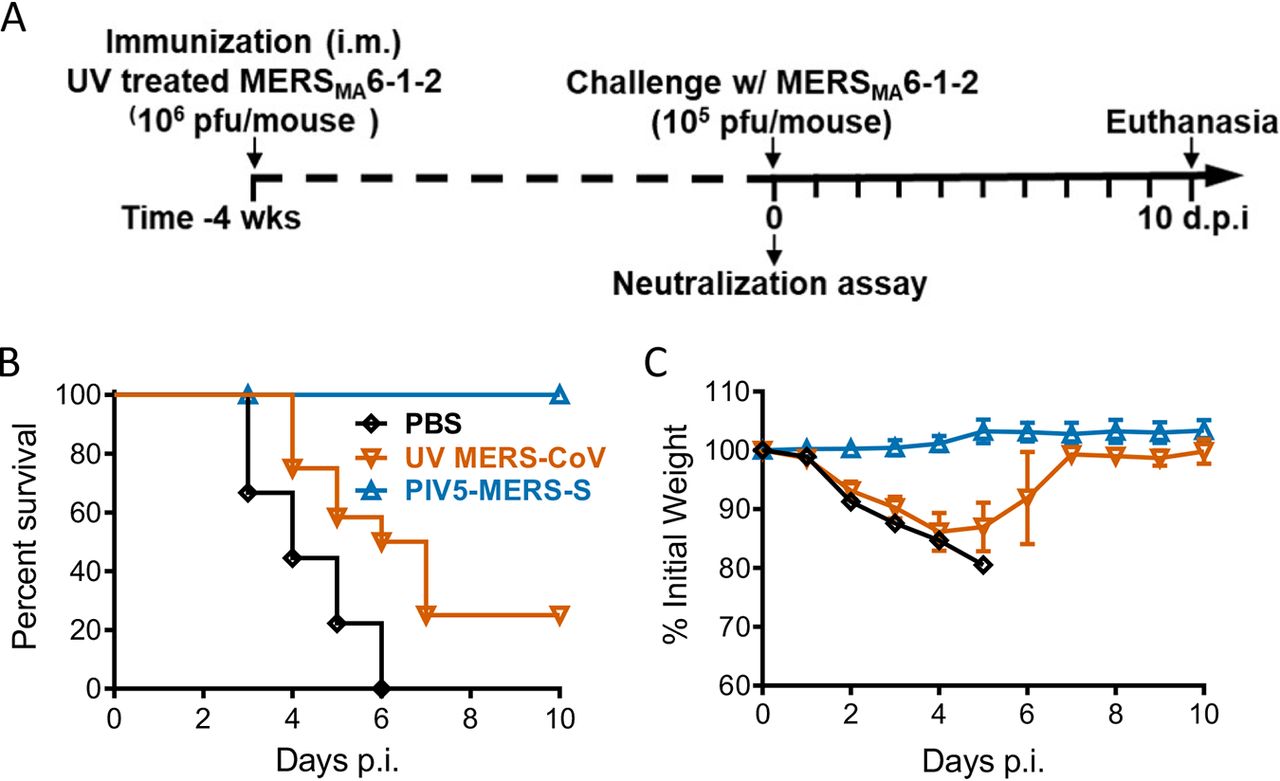
Изображение №6
Далее было проведено исследование легких мышей, которые были сначала иммунизированы, а затем заражены MERS-CoV (изображение ниже).
Изображение №7
Наблюдалось больше эозинофилов (разновидность лейкоцитов) в легких у мышей, иммунизированных инактивированной MERS-CoV, чем у мышей, иммунизированных PBS или PIV5-MERS-S, после заражения MERSCoV.
По сравнению с PBS периваскулярная эозинофильная инфильтрация была значительно увеличена в инактивированной группе MERS-CoV, однако статистическая разница не наблюдалась по сравнению с группой, иммунизированной PIV5-MERS-S.
Также была проведена оценка образования гиалиновой мембраны в альвеолах легких, что является явным признаком тяжелого легочного заболевания. По сравнению с группой PBS мыши, иммунизированные PIV5-MERS-S, продемонстрировали значительную защиту от образования гиалиновой мембраны. А вот мыши, иммунизированные инактивированным MERS-CoV, показали лишь незначительное снижение образования гиалиновой мембраны, что свидетельствует о низкой эффективности этого метода иммунизации, в сравнении с PIV5-MERS-S.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
В данном труде ученым удалось продемонстрировать вполне успешный метод иммунизации мышей от вируса MERS-CoV, который свободно гуляет по нашей планете с 2012 года.
Основной отличительной чертой разработанного метода является использование другого вируса, а именно ПГ-5, в качестве вирусного вектора. Другими словами, клин клином вышибают.
Использование вируса в качестве транспорта для доставки генов, необходимых для иммунизации, можно считать достаточно необычным методом, однако именно он может способствовать более эффективной борьбе с вирусами. Сами исследователи заявляют, что их труд нацелен не только на борьбу с MERS-CoV, но и с SARS-CoV-2 (COVID-19). Оба вируса принадлежат к одному подсемейству, имеют схожие пути инфицирования и воздействия на клетки-мишени. Если разработанный метод иммунизации доработать, то, возможно, получится создать достаточно действенную вакцину.
Именно этим и планируют заняться авторы сего труда в будущем. Получится ли у них создать супервакцину или нет — покажет время. Однако уже сейчас они достойны всяческих похвал как минимум за то, что обратили внимание на вирус, про который будто все забыли.
Как бы то ни было, нам остается надеяться, что наука и все к ней причастные, как и много раз до этого, смогут побороть все предубеждения и страхи, преодолеть все препятствия и показать, что аналитическое мышление, основанное на знаниях и логике, всегда способно найти выход из любой сложившейся ситуации, какой бы страшной на первый взгляд она ни была.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята.
Немного рекламы 🙂
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Похожие статьи
 Три секретами европейских валют
Три секретами европейских валют Видеосвязь больше не гарантия: как дипфейки подрывают доверие в бизнесе
Видеосвязь больше не гарантия: как дипфейки подрывают доверие в бизнесе Как мыслит языковая модель
Как мыслит языковая модель Обычные телескопы способны обнаружить межзвездные лазерные маяки внеземных цивилизаций
Обычные телескопы способны обнаружить межзвездные лазерные маяки внеземных цивилизаций Здравый смысл против релятивизма
Здравый смысл против релятивизма Анатомия кожи: как работает ее естественная защита?
Анатомия кожи: как работает ее естественная защита? Глобальная Управляющая Система: интерпретация высшего разума через призму ТАУ и кибернетики
Глобальная Управляющая Система: интерпретация высшего разума через призму ТАУ и кибернетики Продолжение обзора самых безумных карманных гаджетов наших дней
Продолжение обзора самых безумных карманных гаджетов наших дней