
Как при помощи генных сетей превратить яйцо сначала в личинку, а затем во взрослую муху? Муха, как и другие насекомые, состоит из сегментов. Сегменты объединяются в три группы — голова, грудь и брюшко. На сегменте может быть пара конечностей. В груди три сегмента, на каждом по паре ножек. Второй грудной сегмент имеет еще крылья, а третий — жужжальца (маленькие рулевые крылышки, практически незаметные без лупы). Брюшко состоит из восьми сегментов, конечностей на них нет. В голове исходно было шесть сегментов, но в ходе эволюции насекомых границы между ними совершенно стерлись. Их конечности — это усики, которыми муха нюхает и щупает, и три пары ротовых конечностей. У жука или таракана это были бы жвалы, максиллы и нижняя губа, а у мухи они все сливаются в хоботок.
Чтобы все органы были на своем месте, каждая клетка должна знать, в каком месте зародыша она находится. Поэтому на ранних стадиях развития зародыша, когда в нем еще нет никаких органов и все клетки выглядят одинаково, в нем появляется биохимическая «координатная сетка» с передне-задней, спинно-брюшной и лево-правой осями. Мутации генов, образующих координатную сетку, могут легко и быстро изменить облик животного — передвинуть органы на другое место, изменить их количество и так далее. Поэтому такие гены важны для эволюции животных и очень активно изучается. Генная сеть разметки зародыша у мухи дрозофилы — любимой игрушки генетиков — пожалуй, самая изученная среди генных сетей животных.
Алгоритм разметки яйца по передне-задней оси в принципе несложный:
1) определить, какой конец яйца будет головой, какой хвостом
2) разделить зародыш вдоль передне-задней оси на 17 сегментов
3) разделить каждый сегмент на переднюю, среднюю и заднюю часть
4) разметить голову (6 сегментов), грудь (3 сегмента) и брюшко (8 сегментов)
5) разметить индивидуальные различия сегментов
Генная сеть, которая этим занимается, имеет модульное устройство. Каждый модуль выполняет свой шаг общего алгоритма. Муха дрозофила оказалась очень удобным объектом для изучения генных сетей развития, потому что у нее встречаются очень яркие и показательные наследственные уродства. Например, мутация antennapedia, при которой усики превращаются в ноги, или ВРОТМНЕНОГИ proboscipedia, когда в ноги превращается часть хоботка. Мутация bithorax дает четырехкрылую муху, у которой кроме обычных крыльев на втором грудном сегменте есть вторая пара крыльев на третьем грудном сегменте.
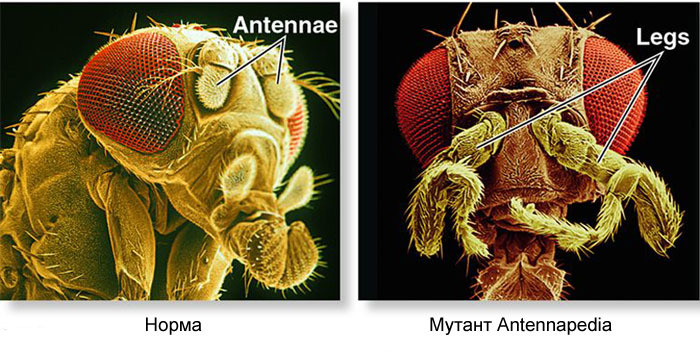

Такие мутации, при которых одна часть тела становится очень похожа на другую, получили название «гомеозисные». Начав с этих мутантов, генетики со временем расшифровали всю сеть генов, управляющих развитием мухи. Эта работа удостоилась Нобелевской премии в 1995 году.
Все начинается с генов полярности яйца — bicoid и nanos. Они работают еще до оплодотворения яйца, пока оно созревает в яичнике мухи-матери. Вспомогательные клетки, накачивающие питательные вещества в яйцо, расположены с одной его стороны, поэтому у яйца с самого начала есть два разных конца. Матричная РНК bicoid накапливается на переднем конце яйца, где она закреплена и не может никуда уплыть. мРНК nanos аналогично закреплена на заднем конце. После оплодотворения по этим мРНК начинают синтезироваться белки. Их молекулы распространяются по яйцу. Поскольку синтез белков bicoid и nanos идет на одном конце яйца, а распад — везде, получается градиент белка bicoid, убывающий от переднего конца зародыша к заднему, и градиент nanos от заднего к переднему концу.
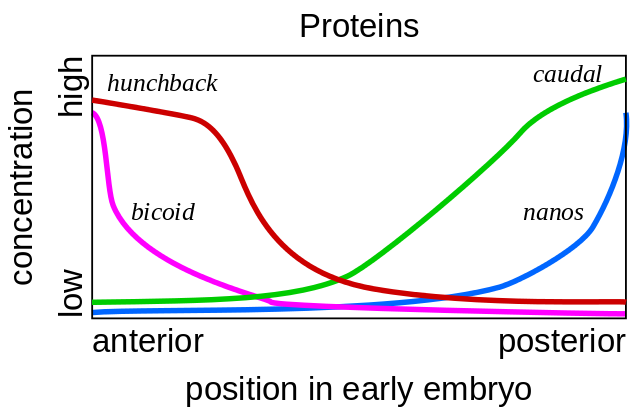
Если микропипеткой ввести мРНК bicoid в другое место яйца, там вырастет вторая голова. Если искусственно выключить мРНК bicoid, получается зародыш без головы и с двумя задницами.
Дальше белки bicoid и nanos включают гены самого зародыша, то есть с них начинает считываться новая мРНК). Первыми начинают работать гены группы gap, от английского gap – промежуток. Первые два гена этой группы — hunchback и caudal – образуют градиенты от головы к хвосту (hunchback) и от хвоста к голове (caudal). Но их градиенты более плавные, чем у bicoid и nanos.
Остальные четыре гена группы gap образуют каждый по одной-две широких (больше одного сегмента) полосы с достаточно четкими границами. Они включаются белками-продуктами вышестоящих генов (bicoid, nanos, hunchback и caudal). Полное выключение одного из генов этой группы дает укороченный зародыш, у которого не хватает нескольких сегментов, тех, в которых в норме этот ген работал.
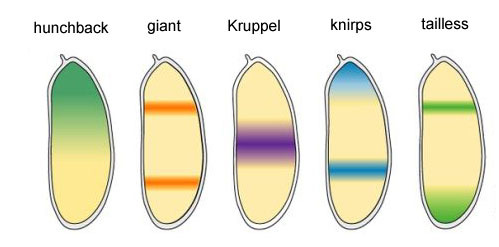
Области активности генов блока gap
Почему получается, что четыре гена блока gap (kruppel, giant, knirps и tailless) делят зародыш на полосы, в каждой из которых работает один из четырех генов?
Во-первых, все они подавляют активность друг друга. В отдельно взятой клетке устойчиво было бы состояние, в котором один ген из такой группы активно работает и подавляет все остальные (триггер). Но развивающееся яйцо мухи устроено сложнее. В нем в этот момент в общей внутриклеточной жидкости (цитоплазме) плавают тысячи отдельных клеточных ядер. Размеры яйца достаточно велики, чтобы скорость распада регуляторных белков была сравнима с их скоростью диффузии на другой конец яйца. При сочетании диффузии и взаимного подавления появляется другое устойчивое состояние, когда яйцо разделено на несколько полос, в каждой из которых работает один ген из группы. Чем медленнее диффузия белков или быстрее распад, тем мельче будут эти полоски или пятна. Участие таких систем (их называют реакционно-диффузионными) в разметке тела животных предсказал еще Алан Тьюринг (да, который придумал «машину Тьюринга») в 1952 году. Тогда мало что было известно про гены и белки, но уравнения, описывающие полосатые и пятнистые структуры, были выведены Тьюрингом верно. Здесь или здесь можно поиграть с простой реакционно-диффузионной моделью из двух веществ и прочувствовать, как она может себя вести.
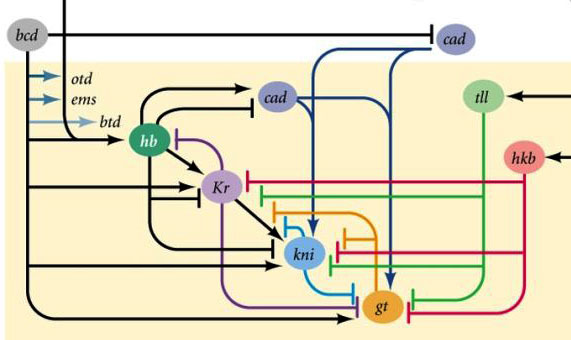
Связи между генами gap и генами полярности яйца. Bcd – bicoid, hb – hunchback, cad – caudal, Kr – Kruppel, kni – knirps, gt – giant, tll – tailless.
Само по себе подавление генами друг друга даст разметку зародыша на какие-то полоски, но расположение полосок активности четырех генов относительно друг друга и концов зародыша будет очень изменчиво. Чтобы полоски были всегда на своем месте (например, для knirps – одна полоска в голове, другая в задней половине между полосками kruppel и giant), нужен второй управляющий сигнал — градиенты bicoid и других белков полярности яйца. Входные функции gap-генов по этим регуляторам имеют максимумы при разных уровнях входного сигнала (такую входную функцию мы вывели в конце второй статьи). Поэтому гены gap включаются каждый своим уровнем белков bicoid, hunchback и caudal, и зоны их активности закономерно расположены от головы до хвоста зародыша.
После того, как гены gap сделали грубую разметку зародыша на 7 полос (они не совпадают ни с сегментами, ни с головой-грудью-брюшком), включается следующий модуль — pair-rule («гены парного правила»). Он делит зародыш уже на сегменты, которых должно быть 17. Модуль pair-rule состоит из пяти генов, и все они, как вы, возможно, догадались, подавляют друг друга. Каждый из этих генов работает либо во всех четных, либо во всех нечетных сегментах — кодируемые ими белки распадаются быстрее, чем белки группы gap, и полоски соответственно получаются более узкие. Выключение части генов этой группы дает укороченный зародыш, у которого исчезают все четные или все нечетные сегменты.
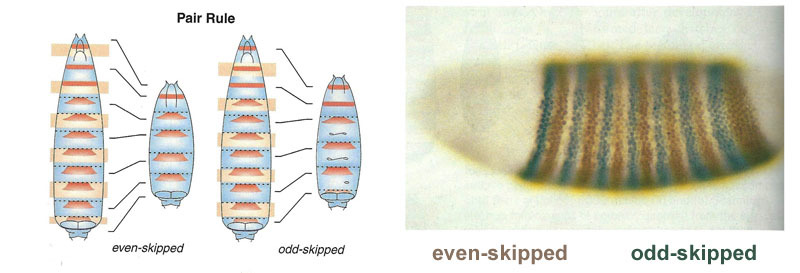
Зоны активности генов из группы pair-rule. На фотографии справа 14 полосок, а не 17, потому что в первых трех сегментах головы работает другая система разметки.
Модуль pair-rule сам по себе, без участия gap и bicoid, может разметить зародыш на узкие полоски, но число этих полосок было бы неустойчиво: сегментов может получиться и 12, и 18, и даже 20. В норме гены pair-rule сложно управляются генами gap, и первая грубая разметка (на 7 зон) помогает точно и надежно сделать окончательную разметку (на 14+3 сегмента).
Следующий пункт алгоритма — разметить переднюю, среднюю и заднюю часть каждого сегмента. Это делают гены segment polarity. Они тоже подавляют друг друга и включаются вышестоящими модулями геной сети (pair-rule и gap). Только параметры диффузии и распада их продуктов подобраны так, что полоски получаются совсем тоненькие — по пол-сегмента. Подавление их друг другом слабее и организовано сложнее, чем у генов pair-rule. Поэтому их полоски активности частично перекрываются и в каждом сегменте получается частая «линейка» из по-разному перекрывающихся полосок.
Выключение генов segment polarity обычно приводят не к укорочению зародыша и потере его частей (как характерно для gap и pair-rule), а к более тонким эффектам. Обычно при этом половина каждого сегмента, в которой должен был работать этот ген, превращается в зеркальное отражение другой половины. Это хорошо заметно, потому что личинка мухи имеет ряды мелких шипиков, направленных назад — для удобства ползания. У мутантов по генам segment polarity появляются ряды шипиков, направленных вперед.
Наконец, наступает очередь самых известных и открытых первыми генов этой сети — модуля hox. Их работа придает сегментам индивидуальность, чтобы на сегментах головы выросли усики и хоботок, а на грудных — ноги и крылья. Гены hox, в отличие от предыдущих модулей, не подавляют активность друг друга и могут работать вместе в одном сегменте зародыша. Странное свойство генов hox, не получившее объяснения до сих пор: они расположены в хромосоме тесной группой, причем в том же порядке, в котором они работают в теле зародыша.
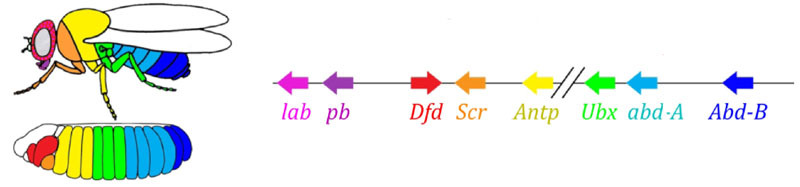
Разметка зародыша устроена по принципу «чем дальше от груди, тем больше hox-генов и активнее работает»: активность гена abdominal-A начинается со 2 сегмента брюшка, нарастает до полной в 4 и продолжается на максимальном уровне во всех последующих сегментах; ген abdominal-B работает с 5 по 8 сегмент брюшка, повышая активность в каждом следующем сегменте. Ген ultrabithorax включается в пол-силы в третьем грудном сегменте и на полную мощность по всему брюшку. При его выключении у мутантов bithorax клетки 3 грудного сегмента не знают, что они — 3 грудной сегмент, в них работает тот же набор hox-генов, что и во 2 грудном (т. е. только antennapedia) и развитие 3 грудного сегмента идет по программе 2 грудного — с крыльями вместо жужжалец и мощными летательными мышцами.
Если выключить все hox-гены в отдельном сегменте, он получится уродливым, но будет больше всего похож на 1 грудной, с ногами и без крыльев. Так получается потому, что далекие предки насекомых были похожи на трилобитов или многоножек. У них было много одинаковых сегментов с ногами. В дальнейшем их тело укоротилось, задняя часть потеряла ноги, став брюшком, а несколько передних сегментов присоединились к голове и их ноги стали деталями ротового аппарата (жвалы, максиллы и нижняя губа).
Математические модели генной сети разметки зародышей
В случае лактозного оперона и петель прямой связи у кишечной палочки можно построить достаточно простые математические модели с умеренным числом параметров, которые предскажут поведение генов при любых входных сигналах. Для лактозного оперона достаточно:
— четырех констант связывания (репрессор и активатор с ДНК, лактоза с репрессором, цАМФ с активатором)
— четырех параметров стехиометрии (4 лактозы на 1 репрессор, 2 цАМФ на 1 активатор, по 1 репрессору и активатору на оперон)
— двух параметров регуляции (влияние репрессора и активатора на вероятность посадки РНК-полимеразы)
— базового уровня активности оперона (когда нет ни репрессора, ни активатора)
— транскрипционной задержки
Итого 12 параметров.
Посмотрим теперь на регуляторные области, скажем, Hox-генов. На схеме ниже один из регуляторных блоков гена abdominal-B.

Цветные полоски обозначают участки связывания регуляторного белка. Весь этот блок имеет размер около 1700 нуклеотидов — в 10 раз длиннее, чем регуляторная область лактозного оперона. В нем 39 мест посадки 7 разных регуляторных белков из всех вышестоящих блоков этой генной сети. И таких регуляторных блоков у abdominal-B четыре. Они включаются последовательно с 5 по 8 сегмент брюшка и создают ступенчатое повышение уровня белка abdominal-B в этих сегментах. То есть для матмодели одного этого гена, по-хорошему, нужно более 100 констант связывания, столько же параметров регуляции, и неизвестное количество параметров для описания взаимодействия между разными входами — в худшем случае до полутора тысяч на попарные взаимодействия между всеми участками посадки белков в одном регуляторном блоке. То есть полная модель этой генной сети заведомо будет иметь более тысячи параметров. И в этом основная проблема: выходных сигналов, которые мы можем измерить в эксперименте, не так много — фактически только уровни каждого из 25 белков и их распределение по длине эмбриона. Поэтому никто даже не пытался смоделировать эту сеть с той же детализацией, что лактозный оперон.
Можно пытаться упростить задачу и описывать действие каждого регуляторного белка на ген только тремя параметрами (связывание, стехиометрия и параметр регуляции). Можно подобрать эти параметры (например, при помощи Марковских цепей) так, чтобы модель точно воспроизводила реальные полоски активности генов. Для первых двух блоков сети (гены полярности яйца и гены gap) такие модели даже сделаны (например).
Проблема в том, что разные запуски алгоритма подбора дают разные параметры модели — но все они работают правильно и воспроизводят активность генов в реальном зародыше мухи! У этих моделей есть общие черты (например, все gap-гены подавляют друг друга и активируются материнскими белками), но различий не меньше, чем сходств.
А раз разные попытки моделирования дают разные результаты, вряд ли хоть один из них совпадает с реальностью. Хуже того, такие модели воспрозводят нормальную работу, но сильно промахиваются в предсказании результатов мутаций.
В следующей части поговорим о том, почему эти неприятности с изучением генных сетей неизбежны в биологии (и сеть разметки зародыша дрозофилы еще одна из простейших в изучении!), что с этим можно сделать, и у каких инженеров стоит поучиться биологам.
Источник


