
Муравьи всегда ассоциировались у нас с трудолюбием, организованностью, коллективизмом и, конечно же, монархической системой управления. Быть муравьем сложно независимо от положения в колонии. Рабочие постоянно работают (да, спасибо Кэп), королева постоянно производит на свет новое потомство, а самцы постоянно заняты оплодотворением королевы. Этот процесс настолько утомителен, что они погибают спустя пару часов после соития. Кастовая система муравьиного общества любопытна не только тем, что определяет роль будущего муравья и его профессию, но и пол. В одних колониях королева может производить на свет только самцов, в других — только самок. Ученые из Калифорнийского университета (США) решили разобраться, какие факторы влияют на выбор пола потомства. Как оказалось, к ним относятся не только факторы окружающей среды, но и генетика королевы. Что именно влияет на пол муравьев, какую роль в выборе играет королева, и зачем нужен такой контроль потомства? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
В отличие от детских сказок, статус монарха накладывает на человека немало ограничений, ответственности и забот, связанных с управлением страной. В муравьиной колонии все точно так же. Начинается монаршеское правление будущей королевы задолго до ее рождения и длится вплоть до ее смерти.

Самка муравьев-листорезов в окружении рабочих.
Когда будущая королева подрастает до нужных размеров, она покидает родительское гнездо и отправляется на поиски подходящего места для новой колонии. Во время «брачного полета», когда молодые самки и самцы покидают родительское гнездо, самцы некоторых видов выделяют феромоны, привлекающие будущих королев. Выбрав подходящего партнера, самка спаривается только с ним, накапливая сперму, которую в последствии использует по мере необходимости.

Самец муравьев вида Camponotus pennsylvanicus.
В поисках новой локации матке помогают сильные крылья, которые она себе откусывает, найдя подходящее место. Затем королева закапывается в землю, создает первую камеру муравейника и начинает откладывать яйца. Поскольку помощников у нее нет, матке приходится самостоятельно заботиться о первом выводке, который она кормит слюнным секретом и/или особыми «кормовыми» яйцами. Учитывая очевидную нехватку провианта, первые родившиеся муравьи являются рабочими, которые вскоре приступают к сбору пищи и уходу за следующим выводком королевы. Любопытно и то, что первые рабочие — самки, но на размножение они не способны.
Колония муравьев, которая началась с одной матки и нескольких рабочих.
Разные виды муравьев демонстрируют порой радикально отличные формы существования колонии. У некоторых муравьев может быть несколько королев, у других — вообще ни одной, а размножение возможно за счет того, что рабочие являются гамэргатами, т.е. способны к размножению (вид Pachycondyla berthoudi).
Важнейшим отличием самок и самцов с точки зрения генетики является набор хромосом: у самок их два, у самцов — один, полученный от материнской самки. Следовательно, когда королева производит на свет новое потомство, она может решать, какого пола оно будет. Если яйца будут оплодотворены спермой, которую самка сохранила после спаривания, то на свет появятся самки, если нет — самцы.

Муравей-рабочий вида Anoplolepis gracilipes (жёлтый сумасшедший муравей).
Можно предположить, учитывая все решения, принимаемые королевой касательно будущего колонии, что именно она является центральной особью в обществе муравьев. Однако, хоть муравьи и живут в монархической системе, тут имеются элементы старого-доброго социализма, так как именно рабочие влияют на общую картину. Рабочие кормят личинок, а потому могут влиять на численность колонии (могут кормить, а могут и не кормить). Если в колонии несколько самок, то те, что производят меньше потомства, могут быть убиты рабочими или же отправлены в соседние колонии.
Как ни посмотри, жизнь муравьиного монарха непроста и, к слову, весьма продолжительна. Королевы муравьев живут до 12-20 лет. Рекордсменом является матка вида Lasius niger (колония в лабораторных условиях), прожившая 28 лет.
Итак, вернемся к теме половой динамики колонии муравьев и того, что на нее влияет.
Как правило, инвестиции в потомство женского или мужского пола являются важным компонентом приспособленности* для размножающихся половым путем организмов.
Приспособленность* — способность к размножению особей с определенным генотипом.
Исследования теории полового распределения привели к множеству открытий, в том числе и к таким понятиям как конфликт между родителями и потомством, эволюция кооперативного воспитания и геномный конфликт.
Конфликты между родителями и потомством, как считают ученые, возникает в разделенных популяциях с сильной локальной конкуренцией партнеров и в популяциях с асимметрией между сестрами и братьями.
Учитывая эволюцию сотрудничества, считается, что родительский контроль над соотношением полов способствует поддержанию кооперативного воспитания. К примеру, вид певчих птиц Acrocephalus sechellensis является ярким примером кооперативного воспитания, когда особи-помощники помогают родительской паре ухаживать за птенцами. При этом помощники помогают растить птенцов либо своих же родителей, либо ближайших родственников.

Acrocephalus sechellensis
Ввиду избытка пищи, некоторая часть особей скорее останется на богатой территории в роли помощника, нежели отправится куда-то для размножения. В результате этих факторов, на свет появляется больше самок (90% от общего числа птенцов). В местах, где пищи мало, на свет появляется больше самцов (80% от общего числа птенцов). Предполагается, что пол будущего потомства предопределяется матерью, но конкретный механизм контроля пока неизвестен. Тем не менее такая система, как у Acrocephalus sechellensis, не является универсальной для всех птиц, коим присуще кооперативное воспитание потомства.
Что касается геномного конфликта, то первым эмпирическим примером стали осы вида Nasonia vitripennis, у которых имеется хромосома, искажающая соотношение полов с женского на 100% мужское.

Nasonia vitripennis
Эта хромосома передается через сперматозоиды в оплодотворенные яйцеклетки, где вызывает потерю других родительских наследуемых хромосом и производит потомство исключительно мужского пола.
Дальнейшее изучение этого вопроса показало, что у некоторых видов идет больший уклон в сторону потомства мужского пола, у других — женского.
В системах, где присутствует внутригеномный конфликт, одним из решений является эволюция подавленной рекомбинации для уменьшения частоты вредоносных мультилокусных* генотипов.
Локус* — местоположение определённого гена на генетической или цитогенетической карте хромосомы.
Это иллюстрируется стандартной моделью эволюции половых хромосом, в которой отбор способствует потере рекомбинации между антагонистическим по полу локусом и определяющим пол локусом на одной и той же хромосоме. Это приводит к Y или W хромосоме, которая присутствует исключительно у представителей одного пола. Согласно «принципу редукции», это также должно происходить вокруг локусов, искажающих соотношение полов. В соответствии с этим предсказанием локусы, искажающие соотношение полов, часто встречаются в регионах с низкой рекомбинацией, но конкретных доказательств нет.
Ученые также отмечают, что принцип редукции может способствовать формированию аутосомных супергенов, контролирующих другие сложные признаки, которые включают эпистатические взаимодействия между двумя или более локусами. Было обнаружено, что такие супергены контролируют фенотипы, включая полиморфную окраску крыльев у бабочек, стратегии спаривания у птиц и грибов, самонесовместимость у растений и социальную организацию колоний у муравьев.
Аутосомные супергены, такие как половые хромосомы, вероятно, представляют собой разрешение прошлого внутригеномного конфликта между двумя или более локусами.
Супергены лежат в основе по крайней мере двух независимо возникших случаев социального полиморфизма у муравьев. У вида Solenopsis invicta численность королев в колонии контролируется супергеном, охватывающим большую часть одной хромосомы. У Formica selysi имеется аналогчиный механизм, но ничего общего в генетическом составе между двумя видами обнаружено не было.
В обеих системах гаплотип, связанный с социальной структурой нескольких королев (т.е. полигинией), является эгоистичным* нарушителем передачи.
Эгоистичные ДНК* — паразитические последовательности ДНК, населяющие геном хозяина. Со временем многие копии эгоистичной ДНК инактивируются мутациями и делециями, оставляя остатки ДНК, называемые мусорной.
В 1976 году Роберт Л. Триверс и Хоуп Хейр в своем труде «Haploidploidy and the Evolution of the Social Insect:…» высказали предположение, что конфликт матка-рабочие, который формируется асимметрией родства внутри каждого гнезда, приводит к предвзятым соотношениям полов.
Соотношение полов представляет собой соотношение репродуктивных самок и самцов и не включает рабочих. Поскольку рабочие более связаны со своими сестрами (среднее родство = 0.75), чем со своими братьями (среднее родство = 0.25), в монохромных колониях с одной маткой они должны отдавать предпочтение производству маток, а не самцов.
Авторы сего труда предположили, что интересы рабочих будут преобладать в этих случаях, так как рабочие обеспечивают всю заботу о выводке, и что это приведет к появлению потомства с большим уклоном в сторону самок, а не самцов. Королевы же в равной степени родственны с потомством мужского и женского пола, поэтому в целом они склонны к соотношению полов 1:1.
В колониях с несколькими матками или одной маткой, оплодотворенной несколькими самцами, более низкая степень родства уменьшает конфликт между матками и рабочими, что приводит к более слабому отбору для предвзятого определения пола.
Поразительно, но некоторые виды социальных насекомых демонстрируют почти полную сегрегацию производства самцов и маток на уровне колонии — явление, известное как «разделенное соотношение полов». Такие крайние случаи наблюдались как минимум у 20 различных родов муравьев, ос и пчел.
Рабочие, чье родство сильнее с королевой, будут специализироваться на производстве новых маток. А рабочие, чье родство с королевой ниже среднего по колонии, будут специализироваться на производстве самцов.

Formica glacialis
Несмотря на множество теоретических и эмпирических исследований, полного понимания того, почему в некоторых колониях преобладают самки/самцы (или вовсе присутствуют только самки или самцы) пока нет.
Посему авторы рассматриваемого нами сегодня труда решили провести свое исследование, в котором попытались найти доказательства генетического контроля в распределении полов у муравьев вида Formica glacialis.
Результаты исследования
GWAS соотношения полов
С помощью метода GWAS (genome-wide association studies, т.е. полногеномный поиск ассоциаций) были идентифицированы многочисленные варианты*, связанные с распределением пола колонии в области хромосомы 3, охватывающей 5.5 Мб* (1A).
Вариант* — более точный термин, эквивалентный термину «мутация». Генетический вариант — это участок ДНК, который отличает геном данного индивида от некого усредненного генома всего вида.
Мб* (мегабаза) или миллион пар оснований; Спаренные основания — пара двух азотистых оснований нуклеотидов на комплементарных цепочках нуклеиновых кислот (противоположных ДНК или одинаковых РНК), соединенная с помощью водородных связей.
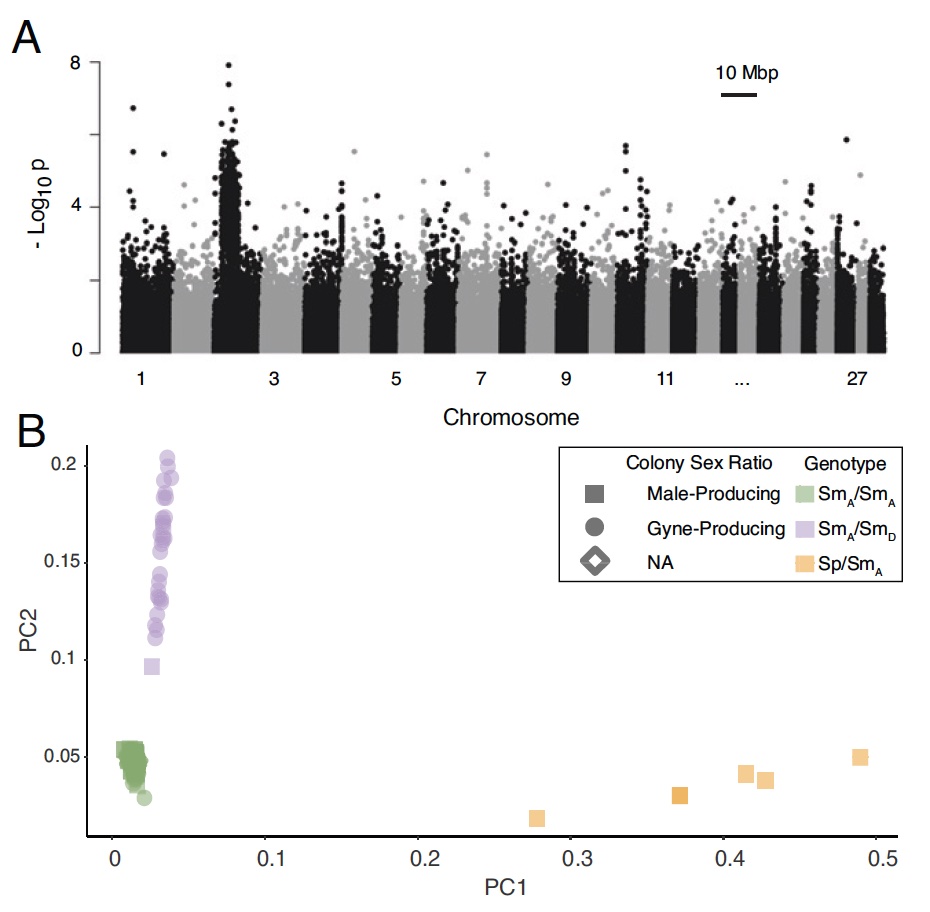
Изображение №1
Анализ вариантов методом PCA (principal component analysis, т.е. анализ главных компонентов) на хромосоме 3 выявил три отдельных кластера генотипов, один из которых наблюдался всего у шести особей (1B).
Среди рабочих с низким значением PC2 (желтые и зеленые кластеры на 1B) 60.2% были собраны из мужских колоний, в то время как 93.3% рабочих с высокими баллами PC2 (фиолетовый кластер на 1B) были из женских колонии.

Изображение №2
Исследование генетической дифференциации (FST) между кластерами генотипов на хромосоме 3 выявило две смежные области: между двумя кластерами с низкими баллами PC1 наблюдалась дифференциация, охватывающая область от 2 до 7.5 Мб (2A), аналогично региону, выявленному в исходном GWAS. Между двумя кластерами с низкими значениями PC2 (оба из которых содержали избыток рабочих из колоний, продуцирующих самцов), была идентифицирована дифференцированная область от примерно 7.5 до 12.5 Мб, а также небольшой пик на 2 Мб (2B).
Связь между социальной структурой и супергеном
Предыдущие исследования показали, что количество маток в колониях вида F. selysi и других европейских видов семейства Formica контролируется социальным супергеном на хромосоме 3. Чтобы определить, является ли суперген на хромосоме 3 аналогичным образом ответственным за число маток в колонии F. glacialis, ученые исследовали генетические вариации еще в 19 дополнительных колониях из других популяций с использованием секвенирования ДНК, ассоциированного с двойным перевариванием рестриктаза (ddRADseq).
Была рассчитана противоположная гомозиготность среди особей внутри одного гнезда (т.е. наличие двух альтернативных гомозиготных генотипов) для определения социальной структуры колонии (3А).

Изображение №3
Для каждой колонии было подсчитано количество локусов, которые показали противоположную гомозиготность в выборке из восьми особей-рабочих из одной колонии.
Для любого единственного двуаллельного SNP не существует комбинации родительских генотипов, которая дала бы оба альтернативных гомозиготных генотипа в наборе множественных родственных потомков (например, рабочих из моногинной* колонии). Напротив, вполне возможно получить оба гомозиготных генотипа в группе рабочих, происходящих от более чем двух родителей, как и ожидалось в полигинной* колонии.
Моногинная колония* — колония с одной маткой.
Полигинная колония* — колония с несколькими матками.
На практике моногинные колонии могут иметь низкие, но ненулевые уровни очевидной противоположной гомозиготности из-за редких ошибок генотипирования или низкие или умеренные уровни в колониях, возглавляемых одной маткой с несколькими оплодотворителями.
В наборе данных, использованном для данного труда, 11 колоний (58%) демонстрировали очень низкие уровни противоположной гомозиготности и были признаны моногинными, в то время как четыре колонии (21%) имели высокие уровни противоположной гомозиготности и были признаны полигинными (3A). Четыре колонии с промежуточными уровнями противоположной гомозиготности были признаны «неопределенными».
Вариация противоположной гомозиготности отображена в области супергена (3B). В частности, однонуклеотидные полиморфизмы (SNP), которые были в значительной степени связаны с вариациями социальной структуры в данных ddRADseq, были локализованы в области от 7.5 до 12.5 Мб (3C), соответствующей области, идентифицированной на 2B.
PCA анализ маркеров на хромосоме 3, которые были общими в наборах данных по полному геному и ddRADseq, показал, что колонии, которые были оценены как полигинные на основе высокой частоты противоположной гомозиготности (красный цвет на 3D), постоянно демонстрировали один генотип. Этот генотип был общим с шестью индивидуумами из библиотеки полногеномного секвенирования, которые сформировали желтый кластер на 1В. Эти особи были гетерозиготными по двум альтернативным супергенным гаплотипам, один из которых, по-видимому, встречается исключительно в полигинных колониях.
Основываясь на результатах ddRADseq, ученые пришли к выводу, что эти три колонии (4% из 71 колонии, собранной для анализа соотношения полов) из целевой популяции, вероятно, будут полигинными. Следуя обозначениям, разработанным для описания гаплотипов, ассоциированных с моногином и полигином у F. selysi (Sm и Sp, где S — суперген, а m и p — социальная форма колонии), ученые определили гаплотип F. glacialis как Sp.
Стоит отметить, что Sp, обнаруженный у других видов Formica, охватывает примерно 10.5 Мб хромосомы 3, от 2 Мб до примерно 12.5 Мб, тогда как Sp, идентифицированный у F. glacialis, был короче.
Основываясь на регионах дифференциации среди кластеров генотипов (изображение №2), можно предположить, что особи из фиолетового кластера несут два альтернативных супергенных гаплотипа в области от 2 до 7.5 Мб хромосомы 3, что впоследствии было подтверждено целевым генотипированием (изображение №4). Один из этих гаплотипов был обнаружен почти исключительно в колониях, продуцирующих самок. Другой гаплотип обычно был гомозиготным в колониях, продуцирующих самцов.
Поскольку один генотип был связан с производством дочерей в моногинных колониях, ученые назвали эти аллели в честь мифических братьев Даная и Эгипта — SmD и SmA соответственно. По легенде, у первого было 50 дочерей, у второго — 50 сыновей.
У особей из колонии, где на свет появлялись самки, был обнаружен генотип SmA/SmD. А у особей из колоний, где рождались самцы, был обнаружен генотип SmA/SmA (отмечено зеленым на 1B).
Распределение генотипов по колониям
Анализ методом PCR-RFLP (polymerase chain reaction restriction fragment length polymorphism) был использован для определения распределения вышеописанных трех генотипов среди большого числа особей из разных колоний и популяций.
Рабочие из колоний самок были смесью гетерозигот SmA/SmD и гомозигот SmA/SmA, в то время как рабочие из колоний самцов чаще всего были гомозиготами SmA/SmA или гетерозиготами Sp/SmA (4A).

Изображение №4
Это предполагает, что колонии, где производятся самки, возглавляются матками SmA/SmD, которые продуцируют смесь самок и рабочих SmA/SmD и SmA/SmA. А вот колонии, где производятся самцы, возглавляются матками SmA/SmA (4B).
В результате подавляющее большинство самцов в популяции должно иметь генотип SmA. Рассмотрение каждой колонии выявило, что 31 из 34 колоний, продуцирующих самок, содержала по крайней мере 1 рабочего SmA / SmD из 8 генотипированных. А 27 из 34 колоний, продуцирующих самцов, содержали только рабочих SmA/SmA и самцов SmA (4С). Среди оставшихся колоний, продуцирующих самцов, три содержали только рабочих Sp/SmA и самцов Sp или самцов SmA (и, вероятно, были полигинными).
Далее ученые вывели генотипы особей из колоний с известной социальной структурой, используя набор диагностических данных SNP. В этих дополнительных популяциях было выявлено, что две моногинные колонии содержали исключительно рабочих SmA/SmA, тогда как девять содержали смесь рабочих SmA/SmD и SmA/SmA (4D). Все четыре полигинные колонии содержали рабочих Sp/SmA. А в одной колонии был выявлен лишь один Sp/SmD рабочий.

Formica podzolica
В дополнение к основному исследованию, которое было нацелено на изучение вида F. glacialis, ученые решили рассмотреть вид F. podzolica, являющийся близким родственником первого. В то время как анализ GWAS был безрезультатным, наблюдались аналогичные F. glacialis качественные закономерности в геномной дифференциации между кластерами генотипов, идентифицированными в PCA (изображение №5).

Изображение №5
Индивиды из этих двух PCA кластеров (5A) демонстрируют повышенную генетическую дифференциацию от 2 до 7.5 Мб вдоль хромосомы 3 (5B). Колонии, продуцирующие самок, содержали смесь гетерозигот SmA/SmD и гомозигот SmA/SmA. Большинство колоний, продуцирующих самцов, содержали исключительно рабочих SmA / SmA (5C). Большое количество SNP, различающих гаплотипы SmA и SmD, сохранялось между F. podzolica и F. glacialis (5D).
Вышеописанные результаты описывают суперген, который, по-видимому, оказывает значительное влияние на соотношение полов в потомстве муравьев. Этот суперген соотношения полов тесно связан с ранее описанным супергеном, лежащим в основе численности маток в колонии муравьев семейства Formica.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В рассмотренном нами сегодня исследовании ученые решили выяснить, что является причиной необычного распределения полов внутри популяции колонии муравьев.
Дело в том, что в некоторых колониях можно встретить исключительно самцов, а в других — только самок. Одним из основных факторов, влияющих на это явление оказался суперген, определяющий соотношение полов будущего потомства.
Однако, как отмечают сами авторы труда, генетика не является единственным фактором влияющим на пол колонии. К примеру, матка может принять решение не использовать сохраненную сперму для оплодотворения яиц, в результате чего на свет появятся исключительно самцы. Также есть задокументированные наблюдения того, что количество пищи влияет на пол будущего потомства. Оказывается, если в колонию положить дополнительную пищу, то рождаемость самцов значительно снизится.
В будущем ученые намерены провести еще несколько опытов и наблюдений для выявления того, как условия окружающей среды могут влиять на демографию колонии.
Многие люди крайне отрицательно относятся к насекомым, определяя их всех в одну кучу и именуя их паразитами или вредителями. Конечно, если в доме завелись муравьи — это малоприятное событие. Когда-то, живя в очередной съемной квартире, я обнаружил, что на кухне (где же еще) завелись муравьи. Спустя несколько безрезультатных попыток избавиться от незваных гостей, я просто насыпал в блюдце сахар (разведенный небольшим объемом воды) и поставил его под раковиной, где его никто не будет видеть, и оно не будет мешаться под ногами. В результате муравьи приняли этот знак примирения и перестали ползать по всей кухне, предпочитая организовывать караваны к легкодоступному блюдцу. Друзья посчитали меня безумцем (и они отчасти правы), но могу поклясться, что численность тараканов (куда более вредных соседей, чем муравьи) после этого заметно снизилась. Мораль сей басни такова — польза может быть и от самых маленьких существ.
Благодарю за внимание, оставайтесь любопытствующими и отличных выходных, ребята! 🙂
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?

