
Фото сетчатки в разрезе с электронного микроскопа.
Дорогие читатели, в этой статье о цвете я не буду приводить аналогии с цифровым фотоаппаратом и фотошопом для «лучшего» понимания физиологии зрения, как не делал этого и в прошлой статье «О разрешении нашего зрения». Такой приём, при кажущемся удобстве, только усложнит картину мира и запутает вас. Буду вести рассказ последовательно и в меру сложно.
Предисловие: краткая теория цвета и света.
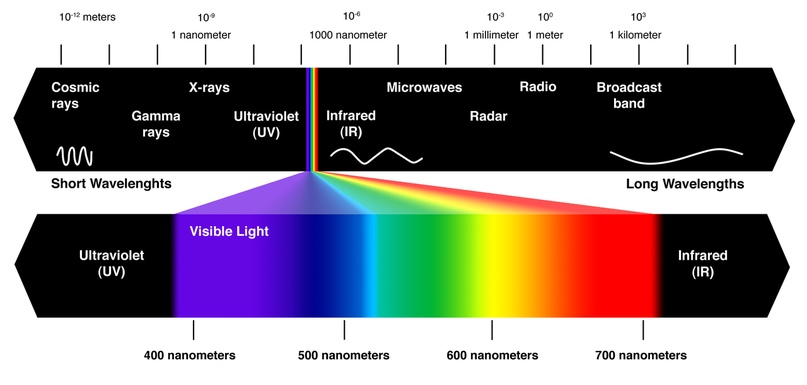
Видимый диапазон.
Свет — это электромагнитные (ЭМ) волны. Из всего разнообразия ЭМ излучения, как видно на картинке выше, наши глаза регистрируют только очень маленькую часть спектра.
Цвет характеризуется тремя величинами:
- — Тон
- — Насыщенность
- — Светлота
Тон — это разные цвета (разные длины волн): синий, красный, зелёный.
Насыщенность: розовый — это ненасыщенный красный.
Светлота: розовый — это светло-красный, а бордовый — тёмно-красный.
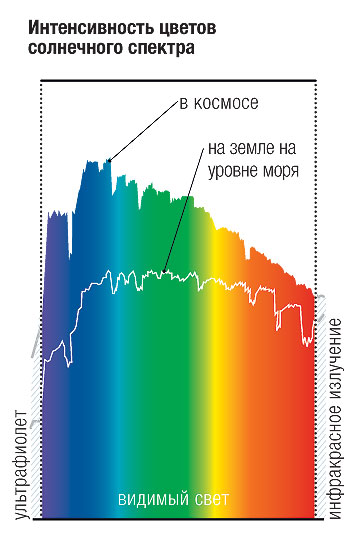
Спектр солнечного света.
Свет от солнца мы видим почти белым с лёгким смещением в жёлтый. Для удобства солнечный свет будем принимать за эталон. На графике выше видно, что атмосфера хорошо поглощает и рассеивает фиолетовую и синюю части спектра (теперь вы знаете, почему небо синее. Для лучшего понимания этого можно почитать про «Рэлеевское рассеяние»).
Почему мы видим зелёные растения зелёными? Потому что они поглощают весь видимый свет, кроме зелёной части, которая отражается и попадает на сетчатку.
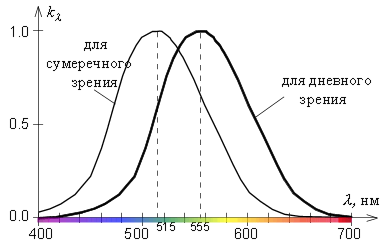
И последний факт перед погружением в физиологию: быстрее всего наша зрительная система реагирует на длину волны света 555 нм — это зелёный цвет с примесью жёлтого. Почему так сложилось? Это вопрос к эволюционной биологии — значит, нашим предкам в какой-то долгий период развития было необходимо хорошо различать этот цвет.
На графике ниже можно увидеть максимум чувствительности для дневного света и для сумерек:
Начнём с общей структуры сетчатки.
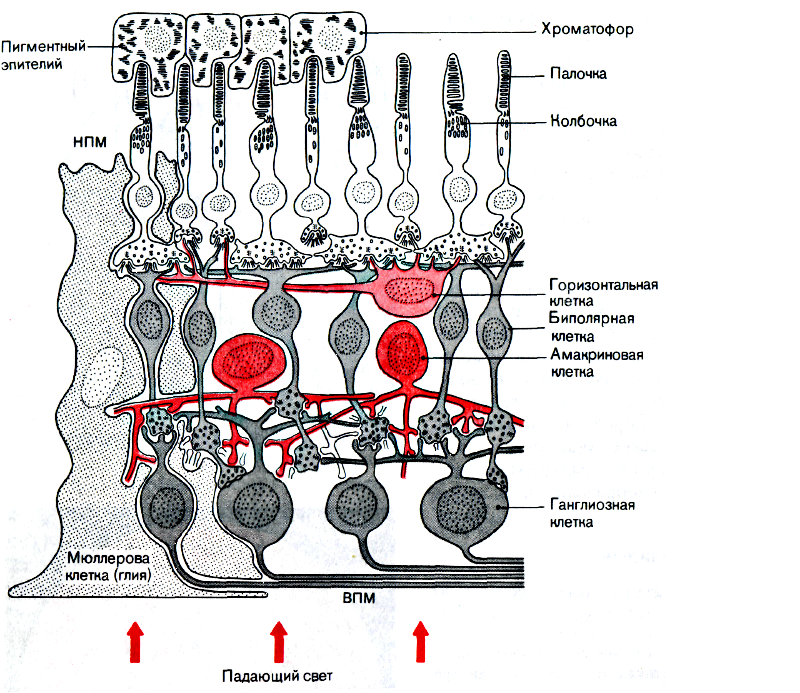
И ещё одна схема для закрепления знаний — всё то же самое, но вдруг кому-то так удобнее:
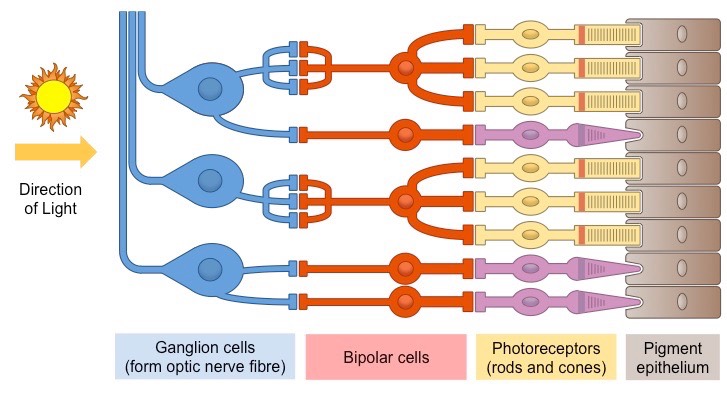
Обратите внимание на красные стрелочки внизу картинки — они указывают путь света сквозь структуры сетчатки. В верхней части схемы показаны рецепторы — палочки и колбочки.
Кого-то из вас может смутить то, что свет попадает сначала на нейроны в сетчатке, а потом на сами рецепторы.
«Как же так? Должно быть наоборот!» — скажете вы. Увы, так «распорядилась» эволюция.
По одной из гипотез, фотороцепторы располагаются ближе к находящемуся сзади пигментному слою, в котором находятся ферменты, участвующие в регенерации фотопигментов.
По другой — нашими очень далёкими предками были ланцетники, чьи глаза находились как бы внутри черепа и улавливали свет сквозь прозрачный скелет, соответственно фоторецепторы были направлены в сторону падающего света. В итоге по ходу всех шагов эволюции сетчатка «не захотела» разворачиваться).
Но не стоит переживать — если вы читаете этот текст и различаете цвета, значит у эволюции всё же получилось) Все слои нейронов сетчатки довольно прозрачны для видимого спектра — этого достаточно, чтобы свет попал на колбочки и палочки с минимальными искажениями.
Итак, сетчатка состоит из трёх типов рецепторов:
- палочки(rods),
- колбочки(cones),
- фоторецепторы(ipRGC).
Палочки содержат пигмент родопсин. Его наибольшая чувствительность находиться в области около 510 нм — бирюзовый цвет.
Колбочки содержат пигмент йодопсин в трёх вариациях. Каждый колбочковый пигмент состоит из хромофора (производное ретинола(витамина А)) и опсина. Хромофор во всех колбочках одинаковый, в то время как опсин разный — это отличие как раз и задаёт разные спектры поглощения!
Немного о видах сигнала.
Ниже показана фотография отдельного фоторецептора, помещённого в сверхтонкую пипетку.
На рецептор направлена полоска монохроматического света. Этот метод позволил измерить мембранный ток фоторецептора.
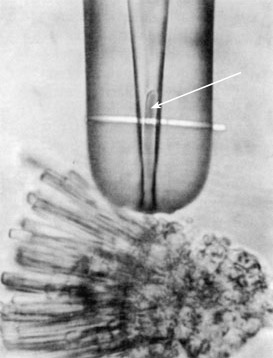
Процесс поглощения фотона и образования сигнала на выходе фоторецептора — фототрансдукция.
При попадании кванта света на фоторецептор в нём происходит распад пигмента и последующий каскад реакций. Рецептор гиперполяризуется от -40мВ до -70мВ. Сигнал на выходе из рецептора не импульсный, а градиентный, т.е. его напряжение зависит от интенсивности света. В результате прекращается передача глутамата от фоторецептора на синапс биполярной клетки и начинается выход нейромедиатора с биполярной на ганглиозную клетку. С ганглиозной клетки выходит импульсный сигнал (потенциал действия ПД), он имеет постоянную амплитуду и длину импульса.
Если на ганглиозную клетку поставить электрод и подключить его к аудио-системе, то при активации этой клетки можно услышать такой сигнал:
youtu.be/ZR7LzRAXNSw?t=94
Пики поглощения колбочек:
— коротковолновые(S) — 426 нм,
— средневолновые(M) — 530 нм,
— длинноволновые(L) — 557 нм.
Обратите внимание на то, что выше указаны именно пики поглощения цвета. Т.е. каждая колбочка реагирует ещё и на соседние цвета с плавным спадом чувствительности.
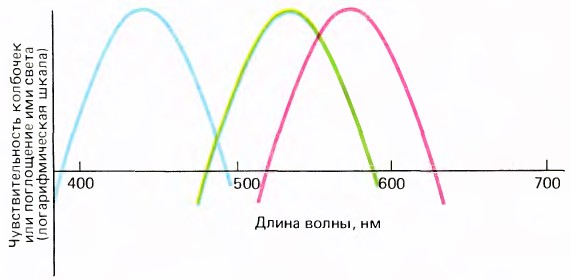
Колбочки имеют широкие зоны чувствительности со значительным перекрыванием. Например, свет с длиной волны 650 нм (красный) вызовет наибольшую реакцию у длинноволновых колбочек и совсем слабый ответ у средневолновых. Т.е. по аналогии — «зелёные» колбочки реагируют не только на зелёный, но и немного на соседние цвета.
Интересный факт, над которым учёные бились почти два столетия — почему при смешении синей и жёлтой красок получается зелёный цвет? Но если взять два источника света, перед одним поставить синее стекло, а перед другим — жёлтое, то в результате смешения получится белый! Этот вопрос удалось решить Герману Гельмгольцу.

Как читать график выше (смешение пигментов)? Очень важно понимать, что жёлтый в данном случае — это не чистый жёлтый с узким спектром в 580 нм, а широкополосный, т.е. это смесь жёлтого с зелёными и красными волнами.
Синий тоже не чистый спектр в 480 нм, а смесь синего с фиолетовым и зелёным.
В результате две смешанные краски или два стекла синего и жёлтого цветов, стоящие друг за другом, поглощают из белого цвета все длины волн, кроме средних — зелёных.
Если же взять монохроматические фильтры на 480 нм и 580 нм и поставить их друг за другом, то сквозь них не пройдёт ничего — не будет перекрытия спектра!
Если осветить белую стену жёлтым и синим фонарями, в результате получится белый цвет. Так происходит по причине «широкополосной» активации колбочек, т.е. всего двумя цветами стимулируются все три типа колбочек и в итоге мы ощущаем белый цвет. По этому принципу работают белые светодиоды — сам излучатель даёт синий цвет, его накрывают жёлтым люминофором — получаем белый свет.
Интересная заметка в книге «Глаз, мозг, зрение» Дэвида Хьюбела на 179 стр.:
«В одной книге, посвященной ткацкому делу, в главе, излагающей теорию цвета, я нашел утверждение, что если вы смешаете в ткани желтые и синие нити, то получите зеленый цвет. На самом же деле получится серый цвет — по биологическим причинам.»
Ещё немного физиологии

Слои нейронов сетчатки:
- Фоторецепторы
- Горизонтальные клетки
- Биполярные клетки
- Амакриновые клетки
- Ганглиозыне клетки
Горизонтальные клетки — отвечают за первичное усиление контраста изображения. Они собирают информацию с нескольких фоторецепторов и образуют первый этап рецептивных полей.
Биполярные клетки — одна из функций этих нейронов — передача сигнала от фоторецепторов к ганглиозным нейронам. Ближе к центру сетчатки один фоторецептор даёт сигнал на один биполяр, дальше от центра происходит конвергенция сигнала, т.е. один биполяр собирает сигнал от множества палочек. Как пример, на периферии зрительного поля на одну такую клетку могут поступать сигналы от 1500 палочек, что позволяет получить хорошую чувствительность зрения при слабом освещении.
Амакриновые клетки — так как на сегодня обнаружено более 33 подтипов данных нейронов, не вижу возможности описать их функции в нескольких абзацах. (Если у кого-то из читателей этой статьи будет свежая информация, то я с удовольствием её добавлю)
Ганглиозные клетки — основная функция — сбор сигнала от предыдущих слоёв нейронов и конвергенция в зрительный нерв. Суммарное количество фоторецепторов сетчатки 100-120 млн — будет превращено в 0,7-1,5 млн нервных волокон в зрительном нерве.
Ещё одна важная функция подтипа ганглиозных клеток ipRGC — регулирование циркадных ритмов в зависимости от яркости освещения и контроль светового рефлекса зрачка.
Теории цветового зрения
Описание теорий сделаю максимально кратким, потому что подробное изложение потянет на отдельную статью. Кому будут нужны подробности — список литературы в конце.
Первую теорию цветового зрения на рубеже 18-19 веков предложили, независимо друг от друга, Джордж Пальмер и Томас Юнг. Она получила название Трихроматическая теория.
Эта теория предполагала наличие трёх типов рецепторов в сетчатке, которые порождают физиологическое ощущение красного, зелёного и синего. Промежуточные оттенки соответственно были истолкованы комбинацией базовых цветов (кардиналов).
Трихроматическая теория очень хорошо объясняет виды цветовой слепоты.
Чтобы понимать механизмы дальтонизма можно прибегнуть к такому эксперименту — предположим, у нас есть пациент страдающий монохромазией (все колбочки в его сетчатке имеют только один пигмент, не важно какой). На сетчатку данного человека посылается поток из 100 фотонов с длиной волны 520 нм (зелёный), а после — 100 фотонов 650 нм (красный). Наш монохромат не получит само ощущение цвета, но сможет отличить эти цвета по их яркости, так как короткие волны обладают большей энергией и их воздействие на фоторецепторы сильнее.
Если же количество длинноволновых фотонов увеличить, чтобы в итоге они вызывали такое же яркостное ощущение как и коротковолновые, то наш больной уже не сможет увидеть различия в источниках света.
Так происходит потому, что фоторецепторы на выходе из сетчатки выдают аналоговый импульсный сигнал — он не способен кодировать информацию о цвете.
Для минимального различия цветовых стимулов в сетчатке должны быть минимум два вида колбочковых пигментов. В этом случае сигнал разных уровней, идущий по разным нервным волокнам, будет в дальнейшем интерпретирован в цвет в зрительной коре.
Так и работают тесты для дальтоников — паттерны изображены разными цветами одинаковой яркости.
Ещё раз про вид сигнала — это аналоговые импульсы, не двоичный код. Сигнал несёт импульсы одинаковой амплитуды, но при этом может изменятся сама частота импульсации — 30 импульсов в секунду или 100.
Трихроматическая теория при всём своём успехе имела ряд недостатков — например, она не могла описать, почему при цветовой слепоте цвета никогда не пропадают единично (только красный или только синий) — хотя по логике самой теории должно быть именно так. А получается попарное выпадение цветов — зелёный вместе с жёлтым или красный и синий.
Примерно в 1870 году на сцену выходит Геринг со своей Опонентной теорией.
Кратко — суть теории в том, что она предлагает четыре базовых цвета, а не три. Эти цвета противоположны (опонентны) друг другу:
- красный/зелёный
- жёлтый/синий
- чёрный/белый
После этих строчек некоторые из вас, кто хорошо шарит в фотошопе, уже начали о чём-то догадываться 😉
Сегодня для описания принята Теория двухэтапного цветового зрения или Теория двойной обработки. Её основоположником был Адольф фон Криз. Но свой финальный вид она обрела в 1957 г. благодаря физиологам Лео Гурвичу и Доротее Джеймсон.
Эта теория объединяет две предыдущих — показывая, что они не противоречат, а дополняют друг друга.
Благодаря развитию методов исследования в физиологии сейчас мы знаем, что первый этап обработки описывается трихроматической теорией, а второй — опонентной.
С развитием молекулярной генетики были установлены пики поглощения для трихроматов:
- короткие волны 426 нм
- средние волны 530 нм
- длинные волны 552 или 557 нм
Что за неразбериха с длинными волнами? Оказывается, возможны генетические различия в формировании пигментов для фоторецепторов, чувствительных к длинным волнам. Это было подтверждено экспериментами по трихроматическому уравниванию.
Да, само ощущение цвета у всех нас немного отличается только по этой причине, но это вариант нормы)
Есть ещё и аномальные трихроматы, у которых имеются все необходимые пигменты, но они синтезируются в сетчатке в совершенно других пропорциях — из-за этого тот цвет, который вы ощущаете как синий, аномальный трихромат может ощущать как красный и есть большая вероятность, что и назовёт он его синим, так как в итоге он имеет все три вида пигментов, позволяющих ему просто различать цвета. Таких аномальных трихроматов можно выявить всё тем же трихроматическим уравниванием.
Подведём итог по теории двухэтапного цветового зрения. Все этапы обработки происходят на уровне сетчатки, прошу не путать с возникновением самого ощущения цвета в отделах зрительной коры.
Порядок таков:
- разделение света на три базовых цвета — Трихроматическая теория
- преобразование трёх цветов в три опонентные пары — Опонентная теория
- интерпретация сигнала в латеральном коленчатом теле ЛКТ
- формирование цветового феномена в зрительной коре
Помню, что обещал не приводить аналогий с программами или устройствами, но на этом моменте придётся согрешить) Теория двухэтапного цветового зрения является полным аналогом конвертации цвета из системы RGB в систему LAB в фотошопе:
- RGB содержит три канала — в каждом по одному цвету
- LAB содержит три канала. Каналы a и b имеют по два цвета, а канал Lightness — чёрный и белый
Схема ниже показывает возможную организацию нейронных сетей при переходе от трихроматического этапа к опонентному:
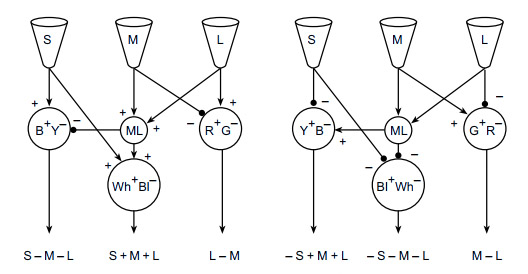
Каков дальнейший путь сигналов из сетчатки после ЛКТ?
До недавнего времени областью зрительной коры, ответственной за распознавание цвета, считалась зона V4.
В 2018 году были проведены исследования по обновлению картирования мозга. Для этого использовались методы объединения данных фМРТ с ретинотопными данными. В результате оказалось, что в коре нет единственного центра, отвечающего за обработку цвета, этим занимаются минимум 5 зон, среди них зона, чувствительная к движению:
- V1
- V8
- VO
- LOC
- MT +
Сами механизмы обработки цвета (формирования цветового ощущения) до сих пор детально не описаны.
Понимаю, что изложение вышло слегка сумбурным, потому что пришлось изучить сотни страниц учебников и исследований. Надеюсь, вам было понятно и интересно 🙂
Литература:
Дэвид Хьюбел — «Глаз, мозг, зрение»
Стивен Палмер — «От фотонов к феноменологии»
Баарс Б., Гейдж Н. — «Мозг, познание, разум»
Джон Николлс, А. Мартин, Б. Валлас, П. Фукс — «От нейрона к мозгу»
Ссылки:
www.ncbi.nlm.nih.gov/pmc/articles/PMC6866375
www.cell.com/fulltext/S0960-9822(08)01433-4
iovs.arvojournals.org/article.aspx?articleid=2161180
en.wikipedia.org/wiki/Fovea_centralis
en.wikipedia.org/wiki/Photoreceptor_cell


